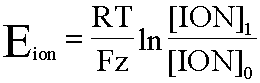VI. RESOLVIENDO EL PROBLEMA
1. La Hipótesis del sodio (Bernstein)
Poco tiempo después que Carlo Matteucci descubrió la corriente de demarcación en el músculo, Du Bois-Reymond confirmó sus hallazgos y además observó que lesionando un segmento del nervio se producía un flujo similar de corriente desde las regiones lesionadas hacia las no-lesionadas (Du Bois Reymond, 1843). Más aún, registró un desplazamiento negativo asociado con la estimulación nerviosa y dijo haber descubierto la identidad entre el principio nervioso (i.e., la esencia de los mensajes nerviosos) y la electricidad.
Du Bois-Reymond no estaba seguro sobre como interpretar los desplazamientos negativos que había descubierto en el músculo. Defendía la idea de que un músculo normal, no dañado, ya tenía una distribución característica de potencial, de manera que la región central era positiva respecto a los extremos tendinosos y la lesión simplemente aumentaba esa diferencia pre-existente de potencial. Además, desarrolló la idea de que las 'partículas electromotrices' debían estar orientadas, o arregladas, de una manera ordenada sobre la superficie del nervio y del músculo, pero no ahondó sobre la naturaleza de esas partículas.
Carl Ludwig (1816-1895) interpretó las ideas de Du Bois-Reymond como equivalentes a comparar los nervios con las líneas telegráficas y las criticó sobre la base de que la resistencia longitudinal de las fibras debía ser muy grande y su aislamiento muy pobre para servir como verdaderos conductores; un argumento perfectamente justificado. De hecho, investigaciones subsecuentes han demostrado que las fibras nerviosas y las musculares pueden ser comparadas con cables submarinos y que estas propiedades son indispensables para la conducción de los impulsos. Sin embargo, las fibras nerviosas son unos cables muy ineficientes, ya que la amplitud de la señal aplicada en un punto disminuye rápidamente con la distancia y el hecho de que sean capaces de trasmitir mensajes por distancias considerables se debe a que son amplificadas frecuentemente durante el viaje, como tuvo que hacerse en el cable submarino.
En 1879 Ludimar Hermann publicó un diagrama con varias hipótesis acerca de la localización de las fuerzas electromotrices en un nervio lesionado y las dos más probables eran: a) la formación de una fuerza electromotriz entre los segmentos lesionados y sanos de la fibra, que llamó 'teoría de la alteración,' y; 2) la existencia de una fuerza electromotriz en la superficie de las fibras sanas, que fue llamada 'teoría de la pre-existencia.'
El principal defensor de la teoría de la 'pre-existencia' fue Bernstein, quien elaboró una hipótesis sobre el papel de la membrana en la generación del impulso nervioso, que descansaba en tres puntos principales:
- todas las células vivas tienen un interior electrolítico rodeado por una membrana permeable a algunos iones;
- a través de la membrana en reposo existe una diferencia de potencial, y;
- durante la actividad la permeabilidad iónica de la membrana disminuye, de manera que la diferencia de potencial se reduce a un valor relativamente bajo.
Inicialmente Bernstein desarrolló el primero de esos tres puntos. En un trabajo publicado en 1868 propuso que una célula viva está rodeada por una membrana que es ligeramente permeable a los iones y que el interior está lleno de un electrolito en el que los iones se mueven libremente, en forma parecida a como lo hacen en el exterior. Posteriormente analizó el segundo punto tomando como base el trabajo de Walter Hermann Nernst (1864-1941) y en 1902 propuso que las membranas del nervio y del músculo se encuentran polarizadas debido a su permeabilidad selectiva al potasio, que se sabía existía en altas concentraciones dentro de la célula. Por lo tanto, si la membrana que rodea las células era permeable a ese catión pero no a los aniones intracelulares, la difusión del potasio debido a su gradiente de concentración tendería a separar las cargas en la membrana y crearía una doble capa, como había sugerido Nernst. De esta forma quedaría explicado el potencial de reposo.
En relación a ese punto, Bernstein pensaba que la excitación era debida a una pérdida transitoria de la permeabilidad selectiva al potasio, lo que anularía temporalmente el potencial de reposo, dando origen a una perturbación propagada o impulso; cuando el exceso de cargas negativas dentro de la célula se uniera a las cargas positivas de afuera, el potencial de difusión tendería a desaparecer. Cuando las investigaciones en electrofisiología fueron enormemente estimuladas por el uso del electrómetro capilar y del galvanómetro de espejo, Bernstein encontró mas apoyo para su teoría, elaborándola con más detalle en su monografía "Elektrobiologie" (1912).
La especulación básica de Bernstein era que las células vivas estaban rodeadas de una membrana que tenía una permeabilidad baja para los iones. Por lo tanto, si se hacía pasar una corriente constante a través de una cámara que contuviera células, la mayor parte de la corriente fluiría alrededor de ellas sin penetrarlas.
Si la membrana era una estructura aislante debería comportarse como un capacitor; esto es, además de su conductancia también tendrá una impedancia. Dado un voltaje aplicado, la cantidad de corriente que cruza la membrana dependerá de su frecuencia, de tal manera que cuando fuera cero (d.c.) fluirá solamente a través de la resistencia; sin embargo, las corrientes alternas serían capaces de cruzar tanto a través del resistor como del capacitor de la membrana. Por lo tanto, midiendo la impedancia de la membrana a diferentes frecuencias sería posible distinguir las contribuciones relativas de la conductancia y de la capacitancia. En los resultados de los experimentos de Höber la impedancia de una masa de eritrocitos era de 1,200 O a 1 KHz, mientras a 10 MHz éste valor decrecía a 200 O. Esto significaba que la resistencia de la suspensión de células estaba localizada en la membrana superficial y no en el interior de las células, confirmando la hipótesis de Bernstein. Esta conclusión fue confirmada experimentalmente por Höber en trabajos publicados en 1910, 1912 y 1913, quien demostró que la membrana de los eritrocitos es un mal conductor, mientras el interior tiene una conductancia del orden de cualquier electrolito biológico. De esta manera, antes de morir Bernstein tuvo la satisfacción de ver confirmada su hipótesis.
Con esto se hizo obvio para los fisiólogos de los primeros años del siglo XX que los iones generan fuerzas electromotrices solamente si se restringe la difusión de una de las especies.
Una base para la teoría moderna del potencial de la membrana es la ecuación derivada a partir de principios físico-químicos por W. Nernst. Esta ecuación define, en términos de las concentraciones de un ion específico a cada lado de la membrana, el potencial al cual no habrá movimiento neto de este ion para ningún lado. En otras palabras, la ecuación define el potencial requerido para oponerse exactamente al movimiento de un ion particular a favor de su gradiente de concentración. Así, si la membrana fuera exclusivamente permeable a un ion específico, entonces el potencial de la membrana sería dado por el potencial de equilibrio a ese ion.
Para cualquier ión la ecuación es,
En esta fórmula,
- Eion, potencial de equilibrio para el ión
- R, constante universal de los gases
- T, temperatura absoluta
- F, constante de Faraday
- z, valencia del ion (sodio, potasio o cloro)
- ln, logaritmo natural
- [ION]i, concentración del ion en el interior de la célula
- [ION]o, concentración del ion en el exterior de la célula
Usando las concentraciones de sodio y potasio medidas en la sangre y axoplasma del calamar, la ecuación de Nernst proporciona valores de -75 mV para el potencial de equilibrio del K+ y +55 mV para el del Na+. Con estos valores Hodgkin y Katz (1949) propusieron que en reposo la membrana es predominantemente permeable al potasio y por lo tanto el potencial de reposo es cercano al EK, pero durante el potencial de acción se hace predominantemente permeable al Na+ y por lo tanto el potencial se mueve hacia el potencial de equilibrio del Na+. En algunos casos, hacia el final del aumento en la permeabilidad al Na+ hay un gran aumento en la permeabilidad al K+, lo que mantiene el potencial cercano al de equilibrio de K+.
La hipótesis de Bernstein predecía un aumento inespecífico de la conductancia de la membrana durante el potencial de acción, con lo que el potencial transmembrana llegaría a cero milivoltios. Sin embargo, las mediciones intracelulares hechas por Hodgkin y Huxley (1939), así como las de Curtis y Cole (1942) en axones gigantes del calamar, mostraron claramente que esa predicción no se cumplía, ya que la amplitud máxima del potencial de acción no era igual al potencial de reposo, sino mucho más grande. El significado de ese 'overshoot' fue aclarado por Hodgkin y Katz (1949), quienes propusieron la 'hipótesis del sodio,' sugiriendo que la despolarización y el 'overshoot' del potencial de acción eran debidos a un gran aumento en la conductancia de la membrana al sodio y un consecuente movimiento de este ión hacia el interior de la célula.
El descubrimiento del sobretiro del potencial de acción, esto es, el hecho de que la polaridad de la membrana se invirtiera (positivo en el interior) durante el valor máximo del potencial de acción, estaba más allá de los fenómenos que podían ser explicados con el concepto de permeabilidad selectiva a los iones potasio que había postulado Bernstein. De hecho, una pérdida transitoria de la permeabilidad al potasio llevaría simplemente a la despolarización de la membrana.
Es curioso que en el mismo volumen de Pflügers Archiv de 1902 en el que apareció el trabajo de Bernstein, también fueron publicadas las bases para la explicación de esas nuevas observaciones. En esta revista se encuentra un trabajo de Overton, en el que se demuestra que la excitabilidad del músculo de rana se pierde si el cloruro de sodio extracelular se reduce a menos del 10% de la concentración normal. Además, la excitación del músculo se acompaña de una pérdida del potasio intracelular y de una ganancia en el sodio intracelular.
Poco después que terminó la Segunda Guerra Mundial, Hodgkin continuó la colaboración que había iniciado con Huxley en 1939. Ellos consideraron el flujo de iones a través de la membrana del nervio durante el potencial de acción en forma cuantitativa y rápidamente llegaron a la conclusión de que para explicar el sobretiro era necesario postular que durante el potencial de acción existía una entrada de iones sodio a la fibra. Así, Hodgkin y Katz decidieron investigar el efecto de la concentración externa de sodio sobre el potencial de acción registrado intracelularmente en axones de la pata el cangrejo y en poco tiempo obtuvieron la mayor parte de los resultados experimentales contenidos en un trabajo publicado en 1949.
Hodgkin colaboró con Huxley para desarrollar una hipótesis que explicaba los efectos del sodio sobre el potencial de acción. Comparando la composición iónica del axoplasma con la del agua de mar se demostraba que la concentración interna de K es aproximadamente 40 veces mayor, mientras la del cloro y del sodio eran aproximadamente 1/10 de las del agua de mar. Por lo tanto, la enorme inversión en el potencial de membrana que ocurre durante el potencial de acción podía ser explicada suponiendo que la membrana del axón no pierde su permeabilidad selectiva al potasio, sino que se vuelve específica para el sodio. Si la concentración interna de sodio era 1/10 de la concentración externa, esto llevaría a una inversión de aproximadamente 60 mV.
Las dos consecuencias principales de esa hipótesis eran: 1) la amplitud del potencial de acción debería depender de la concentración externa de sodio, y; 2) la magnitud de la inversión del potencial de acción, el 'overshoot' (sobretiro), debía aumentar si la concentración externa de sodio aumentaba. Hodgkin y Katz pudieron confirmar experimentalmente ambas predicciones demostrando que: a) el potencial de acción desaparecía cuando la fibra se encontraba en una solución sin sodio, pero recuperaba su valor inicial cuando se restituía el agua de mar, y; b) la dilución del agua de mar con dextrosa isotónica producía una ligera disminución del potencial de reposo y, además, había una disminución importante y reversible de la magnitud del potencial de acción.
Todas esas observaciones eran consistentes con la hipótesis y confirmaban tanto el trabajo de Overton (1902) como los hallazgos de Katz (1947), quien había mostrado que las fibras nerviosas de Carcinus se bloqueban si el cloruro de sodio en el medio externo disminuía a menos del 10-15% del normal.
Basados en esos resultados, Hodgkin y Huxley desarrollaron un modelo en el que un portador negativamente cargado se combinaría con los iones y los transportaría a través de la membrana. En reposo el portador estaría unido por su negatividad a la superficie externa de la membrana y cuando el potencial de la membrana disminuyera por cualquier razón, el portador quedaría libre para moverse y transportar el sodio. Sin embargo, este mecanismo tuvo que ser revisado y cambiado cuando se obtuvieron los resultados de la aplicación del control de voltaje al axón gigante del calamar.