Decusación
La vía corticoespinal del sistema motor humano se origina en las grandes células de Betz localizadas en la capa 5 del área motora primaria y después de recorrer la cápsula interna y los pedúnculos cerebrales, forman el tracto piramidal. Este tracto se divide en la parte posterior del puente y una fracción de los axones cruzan al lado contralateral, mientras los restantes lo hacen mas abajo, en la médula espinal. Esto es, al final todos los axones originados en las motoneuronas superiores eventualmente terminan en el lado contralateral.
El cruzamiento de la vía piramidal significa que un hemisferio cerebral controla los movimientos del lado contrario del cuerpo, lo que es claramente visto en la clínica cuando un accidente cerebral produce parálisis en el lado opuesto. Sin embargo, podríamos preguntarnos, ¿porqué cruzan al lado contralateral las fibras motoras?, ¿no podrían simplemente descender en el mismo lado en el que son originadas?.
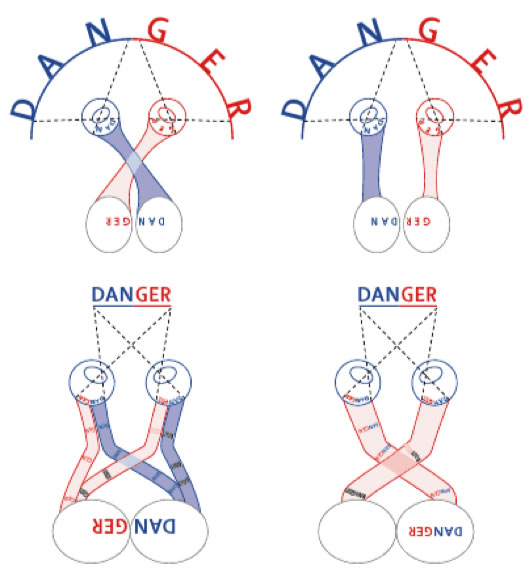 |
Figura 1. Vías visuales cruzadas y no cruzadas. Las imagenes que se forman en la retina son invertidas por el cristalino. Arriba: en animales sin visión estereoscópica (i.e., los campos visuales no se sobreponen), los ojos permiten una vista panorámica para detectar los depredadores. Por lo tanto, un cruzamiento completo de las fibras eferentes retinales en el quiasma es necesario para restaurar una imagen congruente del mundo visual (la palabra "DANGER" es reconstruída, arriba, izquierda). Si no hubiera cruzamiento (arriba, derecha) no se podría identificar la palabra "DANGER" (se vería "GERDAN"). Abajo: en animales que tienen ojos que ven hacia adelante, los campos visuales se sobreponen para permitir la visión estereoscópica, lo que mejora la capacidad motora. Es necesaria una proyección ipsilateral de la mitad temporal de las retinas en el cerebro (abajo, izquierda) para fundir homotópicamente ambas imagenes retinales en el cerebro (abajo, izquierda). Si ocurriera un cruzamiento completo, como en los vertebrados simples, la información estereoscópica estaría comprometida (abajo, derecha), un problema que ocurre en el albinismo humano y otros problemas relacionados. (Vuliemoz et al., 2005)
|
|
Estas preguntas sobre el cruzamiento de fibras también pueden hacerse para cualquier vía que haga lo mismo, como por ejemplo la visual, pero en este caso la repuesta es bastante directa y fue indicada primero por Ramón y Cajal. Cajal analizó el nervio óptico, que después de salir del ojo cruza en parte al lado contralateral en la región llamada quiasma óptico. Aproximadamente el 50% de las fibras cruzan al lado contralateral y después de un análisis morfológico minucioso, llegó a la conclusión de que ese cruzamiento permite que cada hemisferio cerebral obtenga una imagen completa de los objetos que ven los ojos. Esto se demuestra en la Figura 1, donde es claro que la formación de la imagen completa y en la posición adecuada, sólo puede hacerse si las fibras provenientes de la mitad del ojo que detecta un campo visual, se reunen en el hemisferio cerebral con la mitad que proviene de los receptores del ojo opuesto que ven la misma mitad del campo. Esta explicación parece aclarar la razón de la decusación de la vía óptica, pero como no se aplica a la vía motora, la pregunta de porqué se produce la decusación de las pirámides sigue sin respuesta.
Una observación inicial es pertinente. Las fibras del trcto corticoespinal no son las únicas que cruzan al lado opuesto de donde se originan y muchísimas fibras lo hacen también y a varios niveles. Por ejemplo, el cuerpo calloso está formado por axones que cruzan entre los hemisferios cerebrales, pero también hay cruzamiento en varios niveles del cerebro y de la médula espinal. Sin embargo, como en esta sección estamos hablando del control motor, nos centraremos en el tracto corticoespinal y buscaremos una explicación para su cruzamiento.
El cruzamiento de fibras al lado opuesto del cuerpo es una organización que puede haber resultado a partir de una economía en los circuitos sensori-motores y como la decusación es común en todos los vertebrados, empezaremos por examinarla en los vertebrados mas simples y mas antiguos filogenéticamente, los peces.
Historia
Hipócrates (460-380 aC) fue el primero que escribió sobre la naturaleza de las vías motoras, diciendo, "si la lesión está situada en el lado izquierdo [de la cabeza], las convulsiones atacan el lado derecho del cuerpo." Esta información fue refinada 500 años después por Aretaeus de Capadocia (siglo I-II dC), quien notó que cuando las lesiones eran en la cabeza la parálisis era contralateral, pero si las lesiones eran cervicales la parálisis era ipsilateral: "la causa de esto es el intercambio en los origenes de los nervios... cada uno de los cuales pasa al otro lado del de su origen, decusándose en forma parecida a la letra X". Muchos siglos después, en 1710, Pourfour du Petit y Mistichelli identificaron las pirámides en la parte baja del puente como el sitio de la decusación de las fibra motoras y en 1810, Gall y Spürzheim disecaron las fibras motoras desde la decusación piramidal hasta la corteza cerebral y mostraron la continuidad de estas dos estructuras. Así, ya para el siglo XIX y XX, se tenía toda la información sobre el curso y la función del tracto corticoespinal (TCE).
En los humanos el TCE es la mayor vía que media los movimientos voluntarios. Se origina en las neuronas de la capa V de la corteza frontal y parietal (áreas de Brodmann 1-5 y 7) y sólo el 60% de los axones provienen de la corteza motora primaria. El tracto pasa a través de la mitad anterior de la rama posterior de la cápsula interna y antes de llegar al tallo cerebral forma los pedúnculos cerebrales. Durante este trayecto manda colaterales que inervan varias estructuras subcorticales. En la parte baja del bulbo, unos milímetros atrás del cuarto ventrículo, el tracto cruza al lado contralateral formando la decusación de las pirámides, aunque un pequeño porcentaje de fibras (10-25%) permanece en el lado ipsilateral para formar el tracto corticoespinal ventral (o anterior). Las fibras que cruzan forman el tracto corticoespinal lateral, que baja por la médula espinal hasta el último segmento sacro y hacen sinapsis en motoneuronas de la parte dorsolateral del cuerno anterior en varios niveles segmentales, controlando los movimientos finos de la parte distal de las extremidades.
El TCE ventral corre junto a la fisura media anterior de la médula espinal y en los humanos probablemente no se extiende mas allá de la parte superior de la sección torácica. A diferencia del TCE lateral, el tracto ventral muestra una gran arborización distal bilateral, extendiéndose a través de la comisura espinal anterior para inervar interneuronas en ambos cuernos anteriores ventromediales, donde se localizan las motoneuronas que inervan la musculatura axial. Se ha encontrado también un haz corticoespinal que no cruza ventral al TCE lateral.
Evolución del tracto corticoespinal y la decusación motora
Aunque en los humanos el tracto corticoespinal es la vía motora mas importante, es un tracto filogenéticamente jóven que se encuentra sólo en los mamíferos y está particularmente desarrollado en primates y otros pocos animales, aquellos que hacen movimientos precisos con las extremidades. En estas especies, las fibras del TCE son numerosas (hasta un millón), grandes y extensamente mielinizadas para permitir altas velocidades de conducción. Mas importante es que en los primates, los axones corticoespinales generalmente hacen sinápsis directamente en las motoneuronas espinales, mientras que en otros mamíferos las conexiones son principalmente multisinápticas. Esta conectividad uno a uno facilita nuestras excepcionales capacidades motoras, por lo que la interrupción del TCE conlleva problemas persistentes de la capacidad motora fina en los primates, pero el problema es menos pronunciado en las ratas.
Se cree que las variaciones anatómicas del TCE entre las especies y los individuos reflejan su emergencia reciente durante la evolución. A diferencia de los tractos motores polisinápticos, mas primitivos y redundantes, las conexiones de la corteza a las motoneuronas espinales en el TCE son monosinápticas o disinápticas. El TCE se desarrolla tardíamente durante la embriogénesis y en los humanos la maduración se extiende a la vida postnatal, con conexiones y mielinización dentro del primer año de vida. Este desarrollo se refleja en la aparición progresiva de movimientos precisos y la desaparición de los reflejos primitivos (marcha espinal, reflejo de Babinski) en niños en crecimiento. Las proyecciones ipsilaterales del TCE desaparecen durante la niñez.
En los humanos el 75-90% de las fibras del TCE se decusan, pero hay grandes variaciones. En forma típica, la decusación es asimétrica, ya que las fibras que se originan del hemisferio izquierdo cruzan mas extensamente y mas rostralmente que las del hemisferio derecho. Por lo tanto, el lado derecho de la médula espinal es mas grande que el izquierdo, independientemente de la mano usada mas frecuentemente. Unas pocas fibras del TCE pueden cruzar a través del cuerpo calloso y algunas fibras recruzan al lado ipsilateral en la médula espinal. En las ratas el tracto corre en la cadena dorsal, mientras en los humanos, otros primates y gatos, está localizado mas lateralmente en la médula espinal. En algunos insectívoros el tracto que corre en la médula ventral puede dividirse en varios fascículos, mientras en marmotas y topos no hay decusación y en los elefantes y monotremas la decusación se localiza en la protuberancia mas que en el bulbo.
En los vertebrados no-mamíferos, los tractos se originan en el tallo cerebral, igual que los tractos reticuloespinal, vestibuloespinal y rubroespinal, que controlan la mayor parte de las funciones motoras. En contraste con el TCE estos tractos son muy similares en numerosas especies. Los tractos reticuloespinal y vestibuloespinal son las vías mas primitivas y ya se encuentran en los embriones de lampreas (vertebrados primitivos) con proyecciones predominantemente ipsilaterales que controlan la contracción miotomal segmental, aunque también controlan la función de las extremidades. El contacto con las motoneuronas es vía interneuronas.
En los humanos los tractos vestibuloespinal y reticuloespinal controlan el tono muscular, la postura del cuerpo y el balance. El tracto rubroespinal es intermedio entre estos tractos 'primitivos' y el TCE y sólo existe en especies con extremidades o pseudoextremidades, mediando el movimiento de las aletas en las rayas (peces cartilaginosos).
Igual que el TCE, el tracto rubroespinal cruza al lado contralateral en su mayor parte y contiene proyecciones directas a las motoneuronas. Mientras el tracto rubroespinal es muy prominente en los mamíferos cuadrúpedos simples y aún substancial en los primates, regresa en paralelo a la emergencia del TCE y en los humanos consiste en sólo unos pocos cientos de fibras que proyectan exclusivamente a la médula espinal cervical. Paralelo a la regresión de las fibras rubroespinales, las proyecciones rubro-olivares se hicieron mas numerosas. Las fibras que se originan del núcleo rojo fueron incorporándose progresivamente a circuitos cerebro-cerebelo y constituyen la fuente dominante de entradas a las fibras trepadoras. Por lo tanto, mientras que en los humanos el tracto rubroespinal mismo es vestigial, el núcleo rojo aún está involucrado en el control motor, principalmente vía las proyecciones rubro-olivares.
Teleología del cruzamiento en la línea media
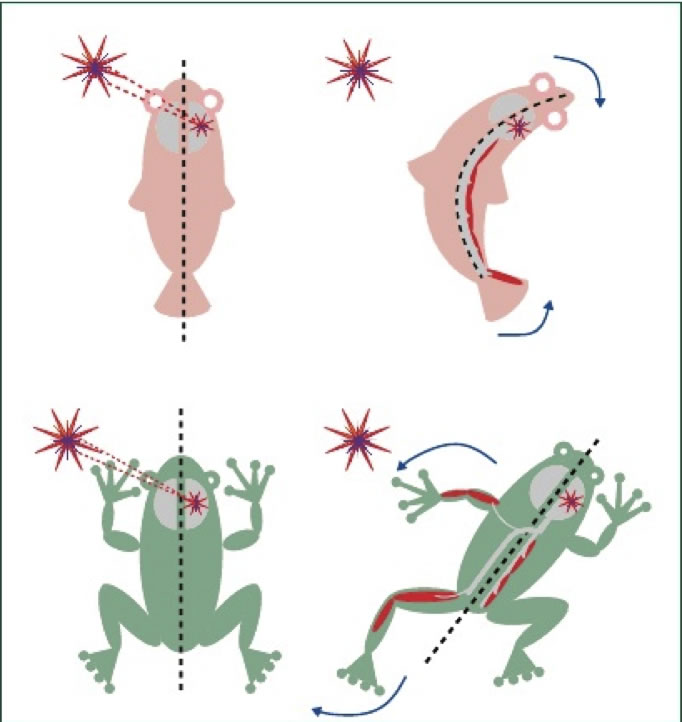 |
| Figura 2. Vías motoras cruzadas y no-cruzadas en el comportamiento de escape. Arriba izquierda. Un estímulo amenazador (estrella) visto en el lado izquierdo activa áreas visuales contralaterales (derecha). Arriba, derecha. Un vertebrado sin extremidades escapa contrayendo la musculatura axial del mismo lado que las áreas visuales activadas, por medio de activación de las vías motoras ipsilaterales (e.g., vía reticuloespinal o vestíbulo espinal). Abajo: un vertebrado con extremidades escapa de un estímulo idéntico extendiendo la musculatura de la extremidad contralateral al área visual activada. Esto ocurre por medio de activación de las vías motoras cruzadas (e.g., rubroespinal o corticoespinal). La contracción de la musculatura axial derecha ocurre como se ve arriba. (Viliemoz et al., 2005). |
|
Ya hace mas de 100 años Santiago Ramón y Cajal se preguntaba sobre la teleología del cruzamiento de las fibras motoras y en un artículo publicado en 1898 proporcionó la explicación mas clara hasta ahora. Sobre la base de que el cristalino invierte las imagenes que se forman en la retina respecto al mundo externo, Cajal indicó que el cruzamiento en el quiasma era necesario para restaurar la continuidad en el cerebro (Figura 2). El cruzamiento es completo o parcial dependiendo de la existencia de visión binocular, la que genera una representación de cada hemisferio visual en el lado opuesto del cerebro. La geometría del tracto visual está dada por las limitaciones ópticas y el cruzamiento de las vías tactíles es necesario para permitir que las dos entradas sensoriales se junten en el cerebro, generando una representación sensorial global contralateral al estímulo. La decusación motora sigue a la representación sensorial cruzada, ya que permite que la extremidad correcta sea activada por la estimulación sensorial. Como la gran dexteridad manual que permite el TCE es muy dependiente de entradas visuales y tactíles adecuadas, la estructura de este tracto es particularmente influída por la anatomía de las vías visuales y sensitivas y hay un extenso cruzamiento. Este cruzamiento generalizado de tractos de entrada y salida ha permitido la obtención de múltiples modalidades sensoriales y motoras, culminando en las grandes áreas de asociación cerebral que son la característica de la corteza humana.
Según el modelo de Cajal, a lo largo de la evolución ha habido una relación inversa entre la proporción de fibras que cruzan en el quiasma y la de fibras del TCE que se decusan. Hasta donde sabemos esto no ha sido probado, pero otro argumento evolutivo también apoya esta hipótesis. Por ejemplo, en especies que no tienen extremidades, como los peces, un estímulo amenazador en el lado izquierdo del cuerpo es percibido por el hemisferio derecho y evoca una reacción de escape por medio de la contracción de la musculatura axial ipsilateral (Figura 2, derecha), que es mediada por los tractos reticuloespinal y vestibuloespinal. En cambio, un vertebrado con extremidades intentará escapar un ataque similar por el lado izquierdo extendiendo las extremidades izquierdas, empujando sobre el suelo para voltear a la derecha. En este caso la respuesta ocurre vía los tractos rubroespinal y corticoespinal, filogenéticamente mas jóvenes, que cruzan la línea media. Aunque Cajal propuso esta hipótesis hace ya mas de 100 años, todavía no ha sido eliminada y es la mejor hasta ahora.
El reflejo inicio-C
Un argumento en favor de la ventaja del cruzamiento de la vía piramidal en un vertebrado simple, el pez, es su reflejo de escape, llamado inicio-C (C-start) y del cual depende su sobrevivencia. Por ejemplo, cuando un depredador se acerca a un pez por su lado izquierdo, la contracción de los músculos debe mover el pez hacia la derecha y alejarlo del peligro, lo que ocurre sólo si se contraen los músculos de la parte derecha del cuerpo. La contracción de los músculos laterales de la mitad derecha del pez hará que primero la cola y después la cabeza se muevan, alejándose del depredador. Una consecuencia de este movimiento es que la forma del pez, vista desde arriba, cambia de una lineal a una convexa hacia el depredador. Esto es, el circuito del reflejo de escape explica la necesidad de la decusación, ya que el reflejo generado en las motoneuronas superiores en un lado del cuerpo tiene que llegar a los músculos en el otro lado del cuerpo.
Referencias
Ramón y Cajal, S. Estructura del quiasma óptico y teoría general de los entrecruzamientos de las vías nerviosas. Rev. Trim. Micrográfica 3: 15–65, 1898.
Vuliemoz, S., O. Raineteau, and D. Jadaubon. Reaching beyond the midline: why are human brains cross wired? Lancet Neurol 4: 87-99, 2005.