
Bernstein como Profesor de Fisiología en la Universidad de Halle. Retrato tomado en 1875. A la derecha está un anuncio de sus conferencias de 1882.
Julius Bernstein nació en Berlín en 1839 en una familia de bien conocidos escritores políticos. Su padre, Aaron Bernstein, era un periodista que en 1845 fundaría la Congregación Berlinesca de la Reforma Judía, y su primo, Eduard Bernstein, era el teórico del Partido Social Demócrata.
Cuando Julius era aún jóven visitó los laboratorios del Instituto de Fisiología de la Universidad de Berlin, dirigido por du Bois-Reymond (1818-1896) y después de estudiar medicina en la Universidad de Breslau (Silesia), regresó a Berlín a trabajar con du Bois-Reymond, haciendo su tesis doctoral sobre la fisiología del músculo de invertebrados. Después pasó siete años como asistente de Hermann von Helmholtz (1821-1894) en Heidelberg.
A los 34 años Bernstein fue nombrado Profesor de Fisiología y Director del Instituto de Fisiología de la Universidad de Halle (cerca de Leipzig), cuando se le tomó la fotografía arriba. Entre 1890 y 1891 fue Rector de esa Universidad y después continuó trabajando ahí hasta su retiro en 1911, muriendo en 1917.
Durante el tiempo que Bernstein trabajó en Heidelberg con Helmholtz, publicó un trabajo en Pflügers Archiv que inició su fama como fisiólogo. Alrededor de 1840 Carlo Matteucci (1811-1868) había observado una corriente que se originaba en el área cortada de un músculo en reposo y fluía a las regiones sin daño, llamándola "corriente de lesión" o "corriente de demarcación". Pocos años después du Bois-Reymond descubrió una disminución temporal en la corriente de lesión, que llamó "variación negativa" y aproximadamente al mismo tiempo, Helmholtz encontró que la velocidad de conducción del "proceso excitatorio" en las preparaciones de nervio-músculo era alrededor de 30 m/seg. Lo que aún se desconocía era si el "proceso excitario" de Helmholtz y la "variación negativa" de Reymond se conducían a la misma velocidad. Hay que recordar que en ese entonces los aparatos de medición eran demasiado lentos para resolver ese problema.
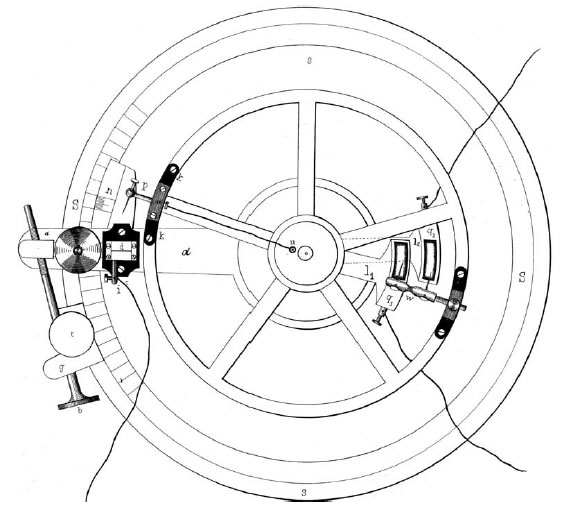
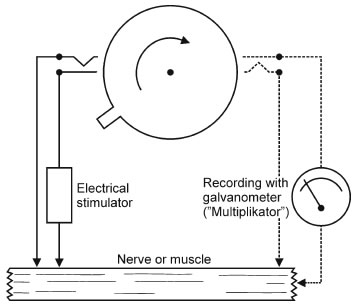
Bernstein resolvió el problema trabajando con un aparato que diseñó y llamó 'reotomo diferencial' o 'cortador de corriente' (Figura 1) y que le permitió determinar el curso temporal de la actividad eléctrica en el nervio y el músculo. El principio del aparato se muestra en la Figura 2. Escencialmente es un "galvanómetro balístico" con un instrumento para medir el tiempo y muestrear. La pieza central es una tornamesa con una pequeña palanca que acciona interruptores que abren y cierran dos circuitos, uno para estimular electricamente un segmento de nervio o músculo, y otro para registrar (muestrear) la "variación negativa" de Reymond con un galvanómetro convencional.
 |
 |
|---|---|
| Figura 1. Vista superior del Reotomo diferencial usado por Bernstein para sus experimentos. | Figura 2. Operación del reotomo diferencial. Primero la palanca de la tornamesa cierra el circuito para estimular y después un segundo interruptor conecta el circuito que registra la actividad del músculo o nervio. |
El interruptor conecta el galvanómetro a los electrodos de registro durante una fracción de milisegundo (generalmente 0.3 mseg), y sólo después de un retardo. El retardo era variable y podía ajustarse con una precisión de milisegundos. Bernstein usó estímulos repetidos y sumó los registros.
En la Figura 3 se muestran las reconstrucciones del curso temporal (arriba) y de la distribución espacial (abajo) de la "variación negativa" en el músculo de rana, conforme es muestreado con el reotomo diferencial. Estos diagramas, publicados en 1868, se consideran la primera descripción precisa del potencial de acción en el nervio.
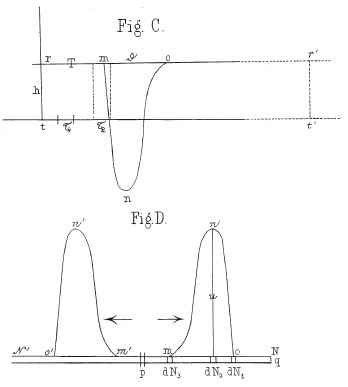
Figura 3. Variación negativa (potencial de acción) de un nervio de rana muestreado con el reotomo diferencial. Arriba. reconstrucción de la señal muestreada y promediada. Abajo. Distribución espacial de la "variación negativa" estimulada en el punto p, moviéndose hacia ambos lados del nervio.
Tres años después Bernstein resumió sus observaciones en una monografía de 240 páginas (Untersuchungen über den Erregungsvorgang im Nerven- und Muskelsysteme) dedicándola a du Bois-Reymond y Helmholtz. Sus principales conclusiones fueron: 1) la "variación negativa" es transitoria; dura 1 mseg y su curso temporal no depende de la fuerza del estímulo; 2) con estímulos fuertes su amplitud excede la de la corriente de lesión ("sobretiro"); por lo tanto, la variación negativa es mas que la 'ruptura' del potencial de reposo. (Después Bernstein escogió ignorar el sobretiro porque no lo veía siempre); 3) la velocidad promedio de propagación era 28.7 m/seg, similar a lo que Helmholtz había medido en 1850. Por lo tanto, supuestamente el "proceso excitatorio" y la "variación negativa" son el mismo fenómeno, y; 4) en el músculo, la variación negativa casi termina antes que se produzca la contracción muscular, lo que parecía indicar que esa variación era un proceso molecular que preparaba la contracción.
La segunda gran contribución de Bernstein a la fisiología fue su teoría de la membrana, publicada en 1902, cuando tenía 64 años de edad. Esta teoría proporcionó la primera explicación fisicoquímica plausible de los eventos bioeléctricos y estaba basada en dos hipótesis que demostró con experimentos relativamente simples.
1. Siguiendo la sugestión de Walter Nernst (1889) sobre los potenciales de difusión de las soluciones químicas a diferentes concentraciones y temperaturas, Bernstein vió los músculos y nervios como cadenas de circuitos de concentración. Los valores de la concentración de las substancias inorgánicas dentro y fuera de la célula ya eran bien conocidos entonces, por lo que Bernstein aplicó las ecuaciones desarrolladas por Nernst para predecir los potenciales eléctricos a partir de los gradientes de concentración y los comparó con la "corriente de lesión o de reposo" que medía en el músculo y en el nervio.
2. Casi al mismo tiempo que Nernst publicaba sus ecuaciones, el físico-químico Wilhelm Ostwald (1853-1932) sugería que el potencial eléctrico a través de membranas artificiales semipermeables se debía a su permeabildad selectiva a los iones. Bernstein aplicó estos conceptos al músculo y nervio y trató la vaina no-conductora como una membrana semipermeable (hay que recordar que entonces no se sabía que las células están rodeadas por una membrana).
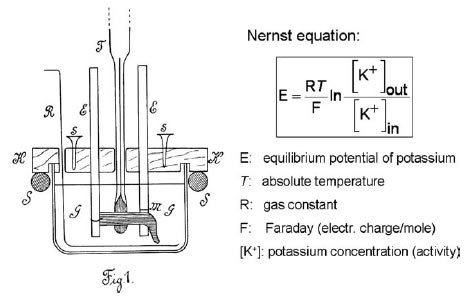
Figura 4. diseño experimental usado por Bernstein para medir el efecto de la temperatura sobre la "corriente de lesión" del músculo y nervio.
El diseño experimental usado por Bernsein para sus mediciones está en la Figura 4. Colocó una preparación aislada de músculo o nervio de rana en aceite en un vaso de vidrio, poniendo al músculo electrodos de yeso mojado en solución salina (no-polarizables) para registro y un termómetro muy sensible. Puso el vaso en un baño de agua, variando la temperatura entre -2 °C y 36 °C y midió cuidadosamente la 'corriente de lesión". Encontró que tanto en el músculo como en el nervio la corriente ("fuerza electromotriz") aumentaba en forma lineal con la temperatura absoluta, resultados experimentales que estaban de acuerdo con las predicciones de la ecuación de Nernst. Esto fue una evidencia de que, "el potencial eléctrico del músculo lesionado es producido por los electrolitos, en particular por sales inorgánicas como K2HPO4, que se encontraban en la fibra muscular sana." (Bernstein, 1902).
En 1912, poco antes de su retiro, Bernstein publicó un resúmen detallado de su trabajo y conceptos en un libro con muchos diagramas. Este libro contiene la primera teoría cuantitativa de la actividad nerviosa y muscular basada en experimentos, mediciones y modelos biofísicos, que finalmente resolvieron el problema planteado por Galvani cuando detectó por primera vez la electricidad animal.