Los movimientos voluntarios son iniciados concientemente en la corteza motora y la corteza de asociación, que planean la actividad, coordinan su secuencia, hacen decisiones sobre las estrategias apropiadas para el comportamiento y evalúan lo correcto de una actividad particular dado el comportamiento corriente o el contexto ambiental, mandando órdenes a los grupos apropiados de motoneuronas en la médula espinal para ejecutar la actividad deseada.
|
|
|
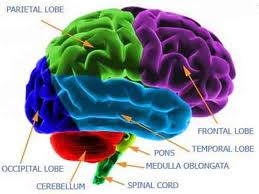
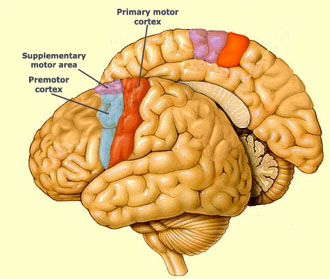
La corteza motora comprende tres áreas diferentes del lóbulo frontal del cerebro, que están inmediatamente anterior al surco central: la corteza motora primaria (área 4 de Brodmann), la corteza premotora y el área motora suplementaria (área 6 de Brodmann) y su estimulación eléctrica inicia movimientos de regiones particulares del cuerpo. La corteza motora primaria (M1) está localizada en la circunvolución precentral y en el lóbulo paracentral anterior de la superficie medial del cerebro y estimulándola con una mínima cantidad de corriente eléctrica, produce movimientos simples de partes individuales del cuerpo. En cambio, la estimulación de la corteza premotora o del área motora suplementaria requiere mayores cantidades de corriente, que resulta en movimientos mas complejos que los producidos por la estimulación de la corteza motora primaria. En monos, la estimulación por periodos prolongados de tiempo (500 mseg) produce el movimiento de una parte del cuerpo, que desde una posición inicial cualquiera, llega a una postura o posición estereotípica. Por lo tanto, parece que la corteza premotora y área motora suplementaria son regiones de nivel superior, que codifican los patrones motores complejos y seleccionan planes motores apropiados para obtener los resultados finales deseados.
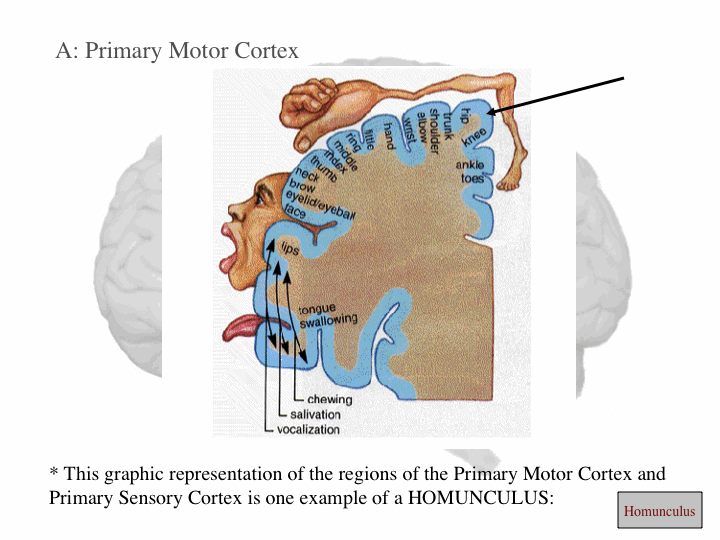
Igual a la corteza somatosensorial de la circunvolución postcentral, la corteza motora primaria está organizada en forma somatotópica y, por ejemplo, la estimulación del lóbulo paracentral anterior inicia movimientos de la pierna contralateral. Si después los electrodos de estimulación se mueven desde una posición dorsomedial a una ventrolateral en la circunvolución precentral, los movimientos producidos progresan desde el torso, brazo y mano, hasta llegar a la cara (la parte mas lateral). La representación de las partes del cuerpo que hacen movimientos precisos y delicados, como las manos y la cara, es desproporcionadamente grande comparada con la de las partes del cuerpo que sólo hacen movimientos gruesos y poco precisos, como el tronco y las piernas, una representación que se denomina 'homúnculo' (Figura 1). La corteza premotora y el área motora suplementaria también contienen mapas somatotópicos.
 |
 |
|---|---|
| Fig. 2. Homúnculo en la corteza motora primaria. | Fig. 3. Modelo tridimensional del homúnculo motor. |
Uno podría predecir que el "homúnculo" se forma porque en la corteza motora el control de los músculos individuales se encuentra en la misma región; esto es, todas las neuronas que controlan el músculo biceps están juntas, todas las neuronas que controlan el músculo triceps también estan juntas y las neuronas que controlan el músculo soleo están juntas en una región un poco mas lejos. Sin embargo, los registros electrofisiológicos han demostrado que eso no es así, ya que los movimientos de los músculos individuales están correlacionados con la actividad de amplias regiones de la corteza motora primaria. En forma similar, la estimulación de regiones estrechas de la corteza motora primaria origina movimientos que requieren la actividad de numerosos músculos. Así, el homúnculo de la corteza motora primaria no representa la actividad de músculos individuales, sino mas bien el movimiento de partes individuales del cuerpo, que frecuentemente requieren la actividad coordinada de grandes grupos de músculos del cuerpo.
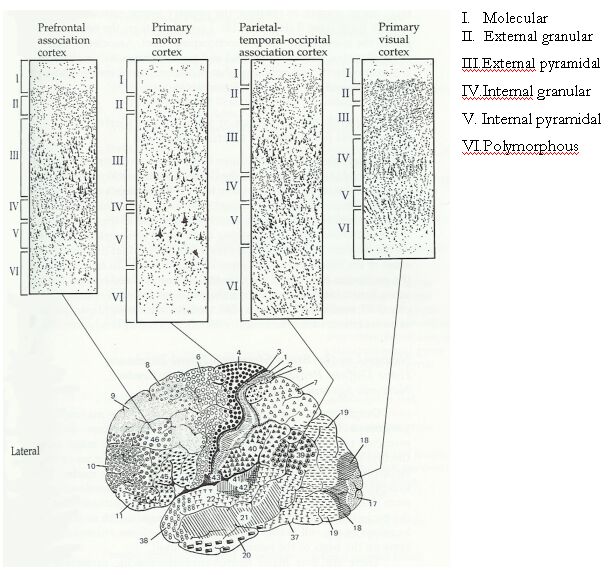
La neocorteza tiene seis capas:
I. Capa molecular. Contiene numerosas dendritas, axones y terminales de axones, pero muy pocos cuerpos celulares.
II. Capa piramidal externa. Contiene células estrelladas muy empacadas, que son interneuronas GABAérgicas.
III. Capa piramidal externa. Contiene células piramidales que usan como trasmisor los aminoácidos excitatorios aspartato y glutamato. Las capas I, II y III conectan regiones corticales adyacentes e integran funciones corticales.
IV. Capa granular interna. Semeja la capa II en que tiene numerosas células estrelladas y recibe la mayor parte de las señales sensoriales.
V. Capa piramidal interna. Semeja la capa III; el tamaño del soma de las neuronas piramidales aumenta de tamaño hacia adentro.
VI. Capa multiforme. Consiste de neuronas fusiformes arregladas perpendicularmente a la superficie cortical.
VII. Contiene una colección de neuronas, axones y dendritas que forman las llamadas columnas corticales.
Como todas las regiones de la neocorteza, la corteza motora primaria tiene seis capas y a diferencia de las áreas sensoriales primarias, es agranular; esto es, no tiene una capa granular con muchas células. En vez de eso, la capa mas distintiva de la corteza motora primaria es la de salidas descendientes (capa 5), que contiene las células gigantes de Betz. Estas células piramidales y otras neuronas de proyección de la corteza motora primaria contribuyen hasta con el 30% de las fibras del tracto corticoespinal. El resto de las fibras provienen de la corteza premotora y el área motora suplementaria (~30%), la corteza somatosensorial (~30%) y la corteza parietal posterior (~10%).
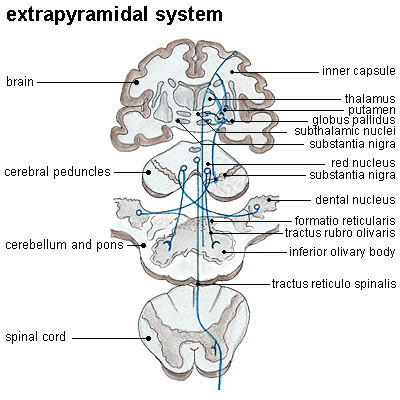
Las clasificación de las vías motoras hecha por Hans Kuypers (1925-1989) está basada en el patrón de terminación de las vías descendentes del cerebro e identifica tres grupos de sistemas. El grupo A lo forma la vía ventromedial del tallo cerebral, que incluye los tractos tectoespinales e interstitioespinal y que se inician en el cerebro medio, los tractos vestibuloespinal lateral y medial, y las proyecciones reticuloespinal y bulboespinal que se inician de la formación reticular del bulbo y la protuberancia. Estas vías descienden en el funiculi ventral y ventrolateral de la médula espinal y terminan en las láminas VII y VIII, la parte ventromedial de la zona intermedia de la substancia gris de la médula espinal, formando un sistema de control para los movimientos de la cabeza, cuello, tronco y la parte proximal de las extremidades; una subdivisión importante de este grupo incluye aquellas vías involucradas en el control de la respiración.
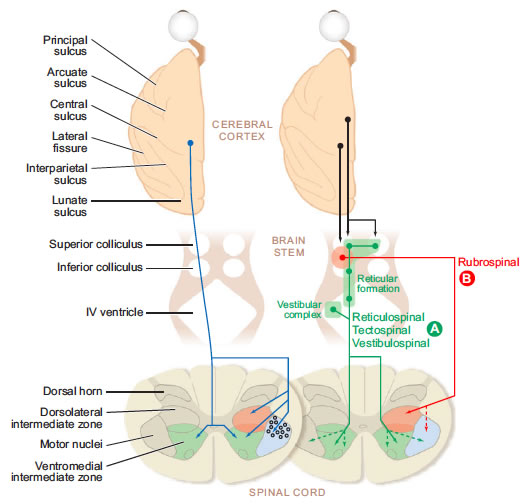
Fig. 5. Distribución de los tractos corticoespinales y las fibras del grupo A (ventromedial) y B (dorsolateral) en el tallo cerebral (Kuypers, 1981).
El grupo B incluye el tracto rubroespinal que se inicia en el núcleo rojo magnocelular y el tracto pontoespinal que se inicia en el tegmentum de la protuberancia ventrolateral. Ambos tractos de caracterizan por su patrón de terminación en las regiones dorsal y lateral de la zona intermedia, dando origen principalmente a neuronas propioceptoras con proyecciones mas locales. Este grupo de vías puede proporcionar capacidad adicional para los movimientos de flexión que involucran los segmentos mas distales de las extremidades, el codo y la muñeca.
Otro grupo de vías (C) influyen sobre funciones motores y otras de la médula espinal y fueron llamadas 'sistema motor emocional'. El componente medial es un sistema difuso de vías que se origina en la parte baja del tallo cerebral (tracto rafeespinal), el tegmentum bulbar ventromedial, el locus coeruleus y el subcoeruleus, y terminan en el cuerno dorsal a todos los niveles espinales. Estas vías usan los neurotrasmisores serotonina y noradrenalina, ejerciendo el primero una gran influencia sobre los reflejos espinales.
Otro componente de este grupo de vías se origina de grupos celulares en el cerebro frontal y medial, y están involucrados en varias actividades motoras específicas que incluyen las reacciones de defensa, dilatación de la pupila, cambios cardiovasculares, vocalización, micturación y conductas sexuales.
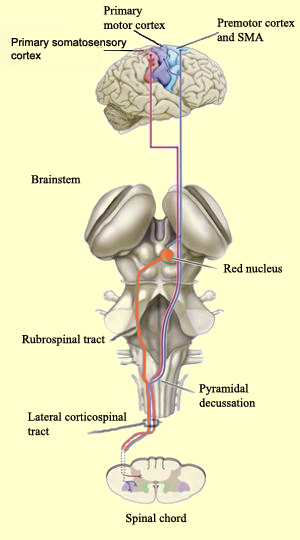
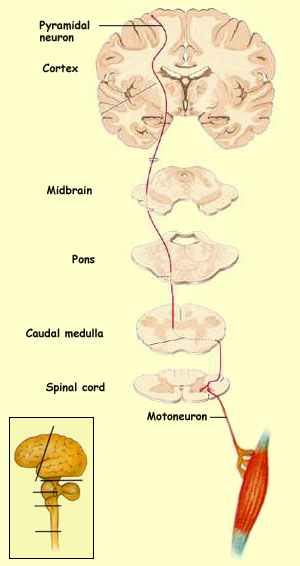
Las vías motoras que se originan en el cerebro o en el tallo cerebral y descienden por la médula espinal hasta las motoneuronas alfa, colocadas en los cuernos ventrales de la médula espinal que, a su vez, mandan sus axones por las raices dorsales para controlar directamente los músculos. Las vías motoras controlan la postura, los reflejos y el tono muscular, así como los movimientos voluntarios concientes. La vía motora mejor conocida es el ‘sistema piramidal', que empieza en las grandes neuronas piramidales de la corteza motora, continúa a lo largo de las pirámides del tallo cerebal y termina en o cerca de las motoneuronas alfa. Este sistema motor es extraordinariamente importante en la clínica porque frecuentemente es afectado por las embolias.
Todas las vías adquieren su nombre por los lugares donde se inician y donde terminan (como el metro), por lo que la vía motora primaria se denomina también corticoespinal, ya que empieza en la corteza y termina en la médula espinal. Específicamente, empieza en la circunvolución precentral, que está inmediatamente anterior al surco central.
La circunvolución precentral tiene varios nombres, como corteza motora primaria, área 4 de Brodman, M1, etc, y los axones de sus neuronas proporcionan la mayor parte del tracto corticoespinal, aunque también contribuyen los axones de neuronas en otras áreas corticales. Una de estas áreas es la 3a, parte de la corteza primaria somatosensorial, que está oculta dentro del surco central.
Si hacemos un corte a través del surco central podemos ver que hay diferencias sutiles entre las circunvoluciones pre y post-central. Aunque todas las áreas de la corteza cerebral tienen seis capas de células, desde la capa I, más superficial y sin células, hasta la capa VI profunda, cada una de ellas tiene componentes celulares ligeramente diferentes y su grosor varía con el área cortical. Así, la circunvolución postcentral, también conocida como corteza somatosensorial primaria, tiene una capa IV muy aparente (en todas las areas corticales la capa IV generalmente recibe información somatosensorial). En cambio, la circunvolución precentral de la corteza motora no recibe muchas entradas sensoriales, por lo que la capa IV no es muy clara, pero como la capa V es responsable de mandar información hacia el tallo cerebral y más abajo, es muy prominente en esta porción de la corteza.
Por otro lado, las vías corticoespinal y corticobulbar están presentes en todos los mamíferos, pero en diferentes extensiones y actúan en paralelo con los sistemas del tallo cerebral. En los animales mas simples (edentados, marsupiales y lagomorfos) la sobreposición en las áreas donde termina el tracto corticoespinal se restringe a las vías rubroespinales. En cambio, en otros mamíferos, que incluyen roedores, carnivoros y primates, hay proyecciones mucho mas extensas del tracto corticoespinal, ya que llegan a todos los niveles de la médula espinal e inervan todas las regiones de la substancia gris, incluyendo las motoneuronas en el cuerno ventral.
 |
 |
 |
Las vías motoras que se originan en el cerebro o el tallo cerebral descienden por la médula espinal hasta las motoneuronas alfa, colocadas en los cuernos ventrales y que mandan sus axones por las raices dorsales para controlar directamente los músculos. Estas vías motoras controlan la postura del cuerpo, los reflejos y el tono muscular, así como los movimientos voluntarios concientes. La vía motora mejor conocida es el ‘sistema piramidal', que empieza en las grandes neuronas piramidales de la corteza motora, continúa a lo largo de las pirámides del tallo cerebral y termina en o cerca de las motoneuronas alfa. Este sistema motor es extraordinariamente importante en la clínica, porque frecuentemente es afectado por accidentes vasculares del cerebro.
El tracto corticoespinal se forma con los axones de las neuronas piramidales de la capa V, principalmente de la corteza motora primaria. En general, entre mas lejos tiene que ir el axón mas grande es la neurona, por lo que en la circunvolución precentral es posible encontrar neuronas especialmente grandes, visibles en el microscopio incluso a poca amplificación. Estas neuronas son llamadas células de Betz y alguna vez se creyó que eran las únicas que originaban el tracto corticoespinal, aunque ahora sabemos que son solamente parte de las células piramidales que forman el tracto. Este tipo de neuronas piramidales se encuentra en todas las áreas de la corteza, porque ‘piramidal’ es solamente una descripción morfológica para neuronas grandes que tienen una forma triangular y apuntan hacia la superficie del cerebro con una larga dendrita apical.
Los axones de las células piramidales entran en la substancia blanca inmediatamente abajo de la capa VI de la corteza, que en todas las circunvoluciones del cerebro contiene los axones que entran o salen. Después, conforme vemos regiones más profundas del cerebro, todas esas rebanadas de substancia blanca se juntan para formar un gran grupo de axones llamado corona radiata, y si vemos aún más profundo en los hemisferios, la corona radiata penetra en los núcleos profundos del cerebro, el caudado y el putamen, dividiéndolos en dos. En este punto el conjunto de todos esos axones se denomina cápsula interna.
La cápsula interna es una gran vía de doble sentido y es muy vulnerable a las embolias. La información sensorial viaja por ella cuando va del tálamo a la corteza y la información motora también viaja por ella en su camino a la médula espinal. En secciones horizontales puede verse que la cápsula interna tiene una rama anterior y una posterior, y tanto la información motora como la somatosensorial viajan en la rama posterior.
Si hacemos cortes horizontales a través del cerebro veremos que alrededor del nivel donde empieza el cerebro medio, la cápsula interna sale de los hemisferios cerebrales uniéndose en un haz. En este punto el grupo de axones forman los llamados pedúnculos cerebrales, que constituyen el piso del cerebro medio y contienen todos los axones descendentes que van al tallo cerebral o a la médula espinal. A diferencia de la cápsula interna, los pedúnculos son vías principalmente de un solo sentido, ya que la mayor parte de los axones se dirigen hacia abajo, mientras los axones sensoriales, ascendentes, toman otras rutas para llegar al tálamo.
Una vez que pasamos el cerebro medio para llegar al puente, dos cosas le pasan a los pedúnculos. Primero, muchos de los axones de la corteza van al puente (las fibras corticopontinas), por lo que ahí se separan para hacer sinápsis. Segundo, el grupo de axones corticoespinales restantes se fragmenta un poco en el puente, de manera que ya no forman un haz compacto y ahora se ven como varios haces de pequeño tamaño. Estas fibras se juntan de nuevo en el bulbo raquídeo para formar las pirámides, que en el tallo humano se ven claramente como dos bordes que corren por la línea media ventral. Las pirámides recorren la longitud completa de la medulla como grandes tractos sobre la superficie ventral.
En el extremo mas caudal del bulbo, cerca del punto donde empezaríamos a llamarla médula espinal cervical, las fibras de las pirámides se cruzan al lado contralateral. Este cruzamiento es llamado ‘decusación de las pirámides’ y se puede identificar porque el surco en la línea media se separa bruscamente de ella, y aunque en un corte transversal parece que solamente un lado cruza, esto es un artefacto, ya que los grandes haces de axones se alternan para cruzar (un poco como los dedos de una mano se pueden interdigitar con los de la otra mano). Así, conforme se hacen secciones más abajo a través de la decusación, la fisura de la línea media se dobla primero hacia un lado y después hacia el otro.
Las pirámides no solamente intercambian posición al decusarse, sino que cada axón que cruza se coloca en la substancia blanca lateral de la médula espinal y cuando se completa la decusación, las fibras corticoespinales ya están en su nueva posición, que ahora se llama tracto corticoespinal y penetra en la substancia gris de la médula espinal. Por ejemplo, las fibras que controlan los músculos de los brazos salen en los niveles cervicales de la médula y hacen sinápsis en el cuerno ventral, ya sea con interneuronas (lo mas común) o directamente con motoneuronas alfa.
En el cuerno anterior de la médula espinal las motoneuronas están organizadas en una forma topográfica de acuerdo a dos reglas, la regla flexor-extensor y la regla distal-proximal. La primera indica que las motoneuronas que inervan los músculos flexores están localizadas en una posición posterior a las motoneuronas que inervan los músculos extensores (músculos de la mano). La segunda dice que las motoneuronas que inervan los músculos distales están localizadas en una posición lateral a las motoneuronas que inervan los músculos proximales (e.g., músculos del tronco).
 |
 |
|---|---|
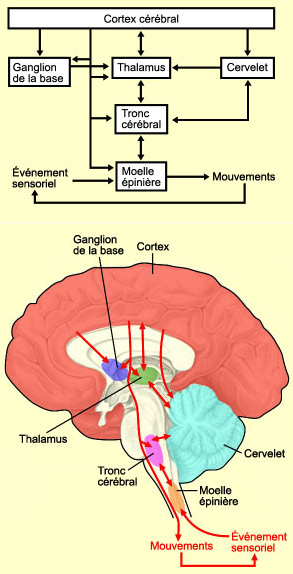
| Figura 7. Vía piramidal para el control del movimiento | Figura 8. Vías extrapiramidales para el control del movimiento |
Generalmente la corteza motora primaria no controla en forma directa músculos individuales, sino que parece controlar movimientos individuales o secuencias de movimientos que requieren la actividad de múltiples grupos musculares. A su vez, las motoneuronas alfa de la médula espinal codifican la fuerza de contracción de grupos de fibras musculares usando un código de frecuencia y el principio del tamaño. Así, de acuerdo con el concepto de organización jerárquica del sistema motor, la información representada por la corteza motora es un nivel de abstracción mayor que la información representada por las motoneuronas espinales.
¿Qué es codificado por las neuronas de la corteza motora primaria? Algunas ideas basadas en datos obtenidos registrando la actividad de estas neuronas en animales de experimentación, conforme la corteza motora primaria codifica los parámetros que definen los movimientos individuales o secuencias simples de movimientos, son las siguientes.
1. Las neuronas de la corteza motora se activan 5-100 mseg antes del inicio de un movimiento; esto es, mas que disparar como resultado de la actividad muscular, estas neuronas están involucradas en mandar comandos a las motoneuronas alfa, que eventualmente harán que los músculos apropiados se contraigan.
2. La corteza motora primaria codifica la fuerza del movimiento, ya que, por ejemplo, la cantidad de fuerza requerida para llevar el brazo de un lugar a otro es mucho mayor si uno tiene en la mano una pelota de boliche, que si uno tiene una pelota de tenis y muchas neuronas en la corteza motora primaria codifican la cantidad de fuerza que es necesaria para hacer ese movimiento. Nótese la diferencia entre la fuerza del movimiento y la fuerza muscular. Mientras una minoría de neuronas de la corteza motora primaria codifican la fuerza muscular individual, un gran número codifica la cantidad de fuerza necesaria para un movimiento particular, independientemente de qué músculos individuales estén siendo ussados. A su vez, bajo el principio del código de frecuencia y el de tamaño, las motoneuronas alfa traducen los comandos de las motoneuronas de la corteza y controlan la cantidad de fuerza generada por los músculos individuales para llevar a cabo el movimiento.
3. La corteza motora primaria codifica la dirección del movimiento. Muchas neuronas en la corteza motora primaria son selectivas para una dirección particular de movimiento; por ejemplo, una célula se activa mas intensamente cuando la mano es movida hacia la izquierda, mientras será inhibida cuando la mano es movida hacia la derecha.
4. La corteza motora primaria codifica la extensión del movimiento. La activación de algunas neuronas está correlacionada con la distancia del movimiento. Por ejemplo, en un mono entrenado para mover el brazo a lugares diferentes que varíen en dirección y distancia del centro, el disparo de muchas neuronas está correlacionado con la dirección del movimiento (como en el punto 3), mientras el disparo de otras neuronas estará correlacionado con la distancia del movimiento. Es mas, algunas neuronas están correlacionadas con la interacción de una distancia y dirección particulares, lo que significa que están correlacionadas con la posición de un blanco particular.
5. Las neuronas de la corteza motora primaria codifican la velocidad del movimiento. Casi todos los movimientos que tienen un blanco siguen una curva de velocidad como función de la distancia que tiene la típica forma de campana. Por ejemplo, cuando la mano mueve un objeto como una taza de café de un lugar a otro (el blanco), se acelera durante la primera mitad del movimiento, llega a un máximo de velocidad aproximadamente a mitad de camino al blanco y después descelera cuando llega al blanco. La frecuencia de disparo de neuronas en la corteza motora primaria de monos que se correlaciona con este perfil de velocidad tiene una forma de campana, demostrando que en estas neuronas la información sobre la velocidad del movimiento está contenida en los trenes de espigas.
La corteza premotora manda axones directamente a la corteza motora primaria y a la médula espinal, haciendo un procesamiento relacionado con el trabajo mas complejo que la corteza motora primaria. La estimulación con un nivel alto de corriente de las áreas premotoras en el mono, produce posturas mas complejas que la estimulación de la corteza motora primaria. Así, parece que la corteza premotora está involucrada en la selección de los planes motores apropiados para los movimientos voluntarios, mientras la corteza motora primaria está involucrada en la ejecución de esos mismos movimientos.
Cuando se entrenan monos para hacer movimientos particulares en respuesta a una señal visual, con un retardo variable entre el inicio de la señal y el inicio del movimiento, los registros de la corteza premotora muestran que muchas neuronas se activan selectivamente durante el intervalo de espera, muchos segundos antes del inicio del movimiento. Por ejemplo, una neurona particular se activa cuando el mono se está preparando para hacer el movimiento hacia la izquierda, pero no lo hará cuando el mono se prepara para hacer el movimento hacia la derecha. Por lo tanto, la activación de la neurona no produce el movimiento mismo, sino que parece estar involucrada en preparar al mono para hacer el movimiento correcto cuando se de la señal. Este tipo de neurona se llama "motoneurona preparada" ("motor-set neuron"), ya que se activa cuando el mono está preparado para hacer un movimiento.
Algunas neuronas premotoras se activan cuando el animal está haciendo una actividad particular, como rompiendo un cacahuate y es interesante que la misma neurona dispara en forma selectiva cuando el animal ve a otro mono o persona romper un cacahuate. La neurona también dispara en forma selectiva al sonido de la cáscara del cacahuate rompiéndose, aún cuando no haya actividad visual o motora. Estas neuronas se denominan "neuronas espejo" porque responden no sólo a una actividad particular del mono, sino también a la vista (o el sonido) de otro individuo haciendo la misma actividad. (Un interesante video sobre neuronas-espejo se encuentra en: http://www.pbs.org/wgbh/nova/sciencenow/3204/01.html)
La corteza premotora ha sido estudiada con MRI funcional cuando una persona observa el video de una mano agarrando un vaso. En una condición el vaso está lleno y rodeado de platos con comida y la implicación es que la persona está agarrando el vaso para beber. En la otra condición el vaso está vacío y rodeado de platos sucios y la implicación es que la persona está agarrando el vaso para limpiar la mesa. En este experimento la corteza premotora está mas activa cuando el sujeto ve el primer video que cuando ve el segundo, aún cuando los movimiento son los mismos. Así, las neuronas de la corteza premotora son sensibles a las intenciones inferidas del movimiento y no sólo al movimiento mismo, como se deduce del contexto conductual en el que ocurre el movimiento.
Varios sujetos fueron estudiados en un experimento con fMRI conforme observan videos de varios actos motores correctos e incorrectos. Una acción correcta es una en la que el movimiento y el objeto asociado son correctos, como ajustar la hora en un reloj. Una acción incorrecta es una en la que la acción es correcta, pero el objeto es incorrecto, como aplicar grasa negra a un zapato café. Un error de movimiento es uno en el que objeto es correcto pero el movimiento es incorrecto, como intentar poner una moneda en un cochinito cuando lestá orientada perpendicular a la ranura. En este experimento la corteza premotora se activa bilateralmente durante los actividades correctas y los intentos de movimientos equivocados; para los errores en el objeto, sólo la corteza premotora del hemisferio izquierdo se activa preferencialmente.
El área motora suplementaria está involucrada en programar secuencias complejas de movimientos y de coordinar movimientos bilaterales. Mientras la corteza premotora parece estar involucrada en seleccionar los programas motores, basados en estímulos visuales o en asociaciones abstractas, el área motora suplementaria parece estar asociada con seleccionar los movimientos, basados en secuencias de movimientos recordados.
Por ejemplo, en un experimento la actividad cerebral fué medida con un barrido PET mientras los sujetos hacían secuencias de movimientos simples y complejos. Cuando los movimientos eran simples, como mover un solo dedo en forma repetida, la corteza motora primaria y la corteza somatosensorial primaria eran activadas en el lado contralateral. Cuando se pedía al sujeto hacer una secuencia compleja de movimientos con los dedos, el AMS era activada bilateralmente, además de la activación de la corteza motora primaria contralateral y la corteza somatosensorial. Finalmente, cuando se pedía al sujeto quedarse quieto, pero practicar mentalmente la secuencia completa de actividades, el AMS se mantenía activa, aunque las áreas de la corteza motora primaria y la corteza somatosensorial quedaban quietas. Por lo tanto, parece que el AMS está involucrada en los movimientos bilaterales y en la práctica mental de estos movimientos.
Los movimientos pueden ser definidos en términos de dinámica (la cantidad de fuerza necesaria para hacer un movimiento) y cinemática (la distancia y ángulos que definen un movimiento particular en el espacio). Muchos planes de movimiento se representan en términos cinemáticos (e.g., mover la mano hacia la izquierda). Sin embargo, para poder instruir a los músculos apropiados a contraerse con la fuerza adecuada, eventualmente el sistema motor debe traducir eso a una representación basada en dinámica. Los registros en monos han mostrado que durante el retardo en el que se prepara a seguir la instrucción de hacer un movimiento, algunas neuronas del AMS cambian su correlación de disparos, desde una representación basada en cinemática a una basada en dinámica, sugiriendo que el AMS tiene un papel fundamental en esta transformación.
El cuarto nivel en la jerarquía motora es la corteza de asociación, en particular la corteza prefrontal y la corteza parietal posterior. Estas áreas cerebrales no son motoras en sentido estricto, ya que su actividad no se correlaciona precisamente con los actos motores individuales, por lo que su estimulación no resulta en una salida motora. Sin embargo, estas áreas son necesarias para asegurar que los movimientos se adapten a las necesidades del organismo y sean apropiados al contexto conductual.
La corteza parietal posterior está involucrada en procesar las relaciones espaciales de los objetos en el mundo y en construir una representación del espacio externo que es independiente de la posición del ojo del observador o de la posición del cuerpo. Esta representación permite una percepción estable del mundo que es independiente de la orientación del observador, así como la representación de las trayectorias deseadas en el espacio que son independientes de la posición del cuerpo. El daño a la corteza parietal posterior puede resultar en varias apraxias, esto es, la incapacidad para hacer movimientos complejos coordinados. Por ejemlo, un paciente con apraxia construccional es incapaz de copiar la configuración de un grupo de bloques en una secuencia apropiada, aunque es capaz de mover con destreza cada bloque individual.
La corteza prefrontal también está involucrada en la evaluación de una actividad particular y los pacientes con daño en ella tienen problemas para procesar la ejecución. Por lo tanto, hacen decisiones conductuales inapropiadas y frecuentemente no pueden anticipar las probables consecuencias de sus acciones, mostranddo además un comportamient impulsivo y frecuentemente son incapaces de rechazar una satisfacción instantánea a cambio de una recompensa mayor a largo plazo.
El sistema motor está organizado en forma jerárquica, pero en vías los diferentes niveles se influyen unos a otros, por lo que el flujo de información es tanto en serie (comunicación entre diferentes niveles) como en paralelo (multiples vías entre cada nivel). Esta organización en paralelo es muy importante, ya que si la organización fuera simplemente en serie, como los eslabones en una cadena, el daño en una parte del sistema produciría parálisis de casi todos los tipos de movimiento. El daño en los niveles superiores resulta en deficits en la planeación, iniciación y coordinación motora, aunque el movimiento aún es posible.
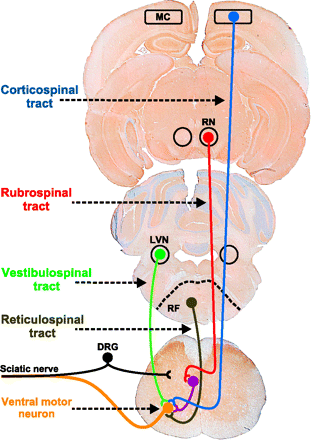
Las vías motoras descendentes están organizadas en dos grandes grupos:
Controlan tanto los músculos proximales como los distales y son responsables de la mayor parte de los movimientos voluntarios de los brazos y las piernas. Incluyen el tracto corticoespinal lateral y el tracto rubroespinal.
Controlan los músculos axiales y son responsables de la postura, balance y control grueso de los músculos axiales y proximales. Incluyen los tractos vestíbulo-espinales (lateral y medial), tractos retículo-espinales (pontino y medular), tracto tecto-espinal y tracto cortico-espinal anterior.
El tracto cortico-espinal se origina en las motoeuronas de la corteza motora y los axones de las motoneuronas de proyección se unen en la capsula interna para viajar a través en los pedúnculos cerebrales en el cerebro medio. A nivel del puente y sobre la superficie ventral del tallo cerebral, estos axones forman las pirámides medulares (por lo que este tracto es llamado piramidal). A nivel de la parte posterior del puente el tracto corticoespinal se divide en dos tractos, uno de ellos comprende aproximadamente el 90% de los axones, que cruzan al lado contralateral en la decusación de las pirámides para formar el tracto corticoespinal lateral. Estos axones continúan a través del funículo lateral de la médula espinal antes de hacer sinápsis, ya sea directamente sobre las motoneuronas alfa o sobre interneuronas en el cuerno ventral de la médla espinal. El resto de los axones (10%) no cruzan en el puente caudal y constituyen el tracto corticoespinal anterior, continuando hacia abajo en el fascículo anterior de la médula espinal. Cuando llegan al segmento espinal en el que terminan, cruzan al lado contralateral a través de la comisura blanca anterior para inervan las motoneuronas o interneuronas en el cuerno anterior. Así, tanto el tracto corticoespinal lateral como el anterior cruzan al lado contralateral, pero lo hacen en diferentes niveles.
El tracto corticoespinal (junto con el tracto corticobulbar) es la vía primaria para los comandos motores de los que depende el movimiento voluntario. El tracto corticoespinal lateral es responsable del control de la musculatura distal y el tracto corticoespinal anterior de la musculatura próximal. Una función particularmente importante es el control fino de los dedos de la mano. El tracto corticoespinal es la única vía descendente en la que algunos axones hacen contacto sináptico directamente sobre las motoneuronas alfa. Supuestamente esta inervación cortical directa es necesaria para permitir que el procesamiento de las poderosas redes de la corteza controlen la actividad de los circuitos espinales que dirigen los movimientos finos de los dedos de la mano.
El porcentaje de axones del tracto corticoespinal que inervan directamente las motoneuronas alfa es mayor en los humanos y primates no humanos que en otros mamíferos, supuestamente reflejando la mayor dexteridad manual de los primates. El daño del tracto corticoespinal resulta en una pérdida permanente del control fino de las extremidades y aunque frecuentemente las vías descendentes paralelas pueden recuperar la función de los movimientos gruesos, no son capaces de generar movimientos finos y precisos. Además del control fino de los músculos distales, el tracto corticoespinal también tiene un papel en el control voluntario de los músculos axiales.
El tracto rubroespinal se origina en el núcleo rojo del cerebro medio. Los axones cruzan inmediatamente al lado contralateral del cerebro, pasando por el tallo cerebral y el funiculus lateral de la médula espinal. Los axones inervan las neuronas espinales en todos los niveles de la médula espinal.
El tracto rubroespinal es una vía alterna a través de la cual los comandos motores voluntarios son mandados a la médula espinal y aunque en muchos animales es una vía grande, en los humanos es relativamente menor. La activación de este tracto produce excitación de los músculos flexores e inhibición de los músculos extensores. Se cree que el tracto rubroespinal tiene un papel en la velocidad del movimiento, ya que sus lesiones resultan en un enlentecimiento temporal del movimiento. Además, como la mayor parte de las entradas del núcleo rojo provienen del cerebelo, probablemente el tracto rubroespinal tiene un papel en la trasmisión de los comandos aprendidos desde el cerebelo a la musculatura. El núcleo rojo recibe también algunas entradas de la corteza motora y por lo tanto, probablemente es una vía importante para la recuperación de alguna función voluntaria después de daño al tracto corticoespinal.
Los dos tractos vestibuloespinales se originan en dos de los cuatro núcleos vestibulares (Figura 2.11). El tracto vestibuloespinal lateral se origina en el núcleo vestibular lateral y pasa a través del tallo cerebral y del funiculus anterior de la médula espinal en el lado ipsilateral, a todos los niveles de la médula espinal. El tracto vestibuloespinal medial se origina en el núcleo vestibular medio, inmediatamente después se divide y pasa bilateralmente a través del tallo cerebral vía el fascículo longitudinal medial y a través del funiculus anterior de la médula espinal, antes de salir en o arriba de la vertebra T6.
Los tractos vestibuloespinales median los ajustes a la postura y los movimientos de la cabeza, ayudando también a mantener el balance. Los movimientos pequeños del cuerpo son detectados por las neuronas sensoriales vestibulares y para contrarrestarlos se mandan comandos a los grupos músculos apropiados de todo el cuerpo a través de los tractos vertibuloespinales. El tracto vestibuloespinal lateral excita los músculos antigravitatorios para ejercer control sobre los cambios posturales necesarios para compensar las inclinaciones del suelo y los movimientos del cuerpo. El tracto vestibuloespinal medial inerva los músculos del cuello para estabilizar la posición de la cabeza conforme nos movemos. También es importante para la coordinación de los movimientos de la cabeza y de los ojos.
Los dos tractos reticuloespinales se originan en la formación reticular del tallo cerebral, que es una gran colección difusa de neuronas organizadas en la protuberancia y el puente. El tracto reticuloespinal pontino se origina en la formación reticular de la protuberancia y se mantiene ipsilateral a través del fasciculo longitudinal medial y el funiculus anterior de la médula espinal, para salir ipsilateralmente en todos los niveles espinales. El tracto reticuloespinal medular se origina en la formación reticular del puente, se mantiene principalmente ipsilateral (aunque algunas fibras cruzan la línea media) a través del funiculus anterior de la médula espinal y sale a todos los niveles.
Los tractos reticulospinales son la mayor alternativa al tracto corticoespinal, por el que las neuronas corticales controlan la función motora debido a sus sinapsis sobre las neuronas reticulares. Estos tractos regulan la sensibilidad de las respuestas flexoras para asegurar que sólo los estímulos dañinos produzcan respuestas. Así, el daño al tracto reticulospinal puede hacer que estímulos no dañinos, como un toque suave, produzcan un reflejo flexor. La formación reticular también contiene circuitos para muchas actividades complejas, como la orientación, el estiramiento y el mantenimiento de una postura compleja. Se cree que también los comandos que inicían los circuitos locomotores en la médula espinal son trasmitidos a través del tracto reticuloespinal medular. Por lo tanto, los tractos reticuloespinales están involucrados en muchos aspectos del control motor, incluyendo la integración de las entradas sensoriales para guiar las salidas motoras.
El tracto tectoespinal se origina en las capas profundas de los colículos superiores e imediatamente después cruza la línea media para pasar anterior al fascículo longitudinal medial y a través de la protuberancia y el puente. Después pasa a través del funiculus anterior de la médula espinal y la mayoría de las fibras terminan en los niveles cervicales superiores.
Se sabe poco sobre la función del tracto tectoespinal, pero como las neuronas del colículo superior (el tectum óptico) reciben entradas provenientes del ojo, se cree que está involucrado en el reflejo que mueve la cabeza para orientarse a los estímulo visuales.
La función mas clara de las vías motoras descendentes es el control de los movimientos voluntarios. Estos movimientos son iniciados en la corteza cerebral y los comandos motores son trasmitidos a la musculatura a través de varias vías descendentes, que incluyen el tracto corticoespinal, el tracto rubroespinal y los tractos reticuloespinales. Ahora, veremos como la corteza motora inicia y coordina los movimientos voluntarios.
Otra función crítica de las vías motoras descendentes es modular los circuitos reflejos en la médula espinal. La adaptatividad de los reflejos espinales puede cambiar dependiendo del contexto conductual, ya que algunas veces la ganancia (fuerza) o incluso el signo (extensión vs flexión) de un reflejo, debe cambiarse para que el movimiento se adapte a las circunstancias. Las vías descendentes son responsables de controlar estas variables. Como ejemplo veamos el reflejo flexor en dos condiciones.
Imaginemos que queremos levantar un plato que está en la parte superior de la estufa y no estamos seguros si está caliente o no. Podriamos intentar tocar ligeramente la superficie y frecuentemente esto bajaría el umbral del reflejo flexor, haciendo mas probable el retirar la mano, aún si el plato no está particularmente caliente (podemos incluso retirar la mano varias veces antes de tocar el plato!). En este caso las vías descendentes han bajado el unbral para producir el reflejo, haciendo fácil que una entrada nociceptiva débil inicie el reflejo; estas vías también pueden cambiar la ganancia del reflejo, haciendo que la respuesta de retirar la mano sea mayor que en general.
Recordemos que en la médula espinal hay dos tipos de motoneuronas: 1) las motoneuronas alfa, que inervan las fibras musculares extrafusales y proporcionan la fuerza de contracción muscular, y; 2) las motoneuronas gama, que inervan las terminales de las fibras intrafusales y ayudan a mantener la tensión de los husos musculares para que sean sensibles a los cambios de longitud en un intervalo grande. La actividad de las motoneuronas alfa y gama debe estar coordinada para que funcionen en una forma congruente, una coordinación que se denomina "coactivación alfa-gama", mientras el ajuste de la sensibilidad del huso muscular por las motoneuronas gama se denomina "bias gama". Ahora, consideremos dos ejemplos:
1. Cuando se da a un músculo la órden de contraerse, los husos musculares se hacen 'guangos', volviéndolos insensibles a nuevos cambios en la longitud del músculo. Para compensar esto, las motoneuronas gama que invervan las fibras musculares intrafusales se activan junto con las motoneuronas alfa, permitiendo que las fibras intrafusales se contraigan con el músculo. Esto mantiene la sensibilidad del músculo para estiramientos inesperados.
2. Cuando un músculo se contrae, el músculo antagonista se alarga durante el movimiento. Un problema obvio ocurre cuando consideramos el reflejo al estiramiento del músculo antagonista. Si la contracción del músculo agonista produce la activación del reflejo al estiramiento del músculo antagonista, este se contraerá para resistir el movimiento de la extremidad. Entonces, ¿cómo es posible flexionar una articulación si el reflejo al estiramiento del músculo extensor trata de extenderlo? El problema se resuelve con la coactivación alfa-gama, que durante un movimiento voluntario relaja la contracción de las fibras intrafusales del músculo antagonista, permitiendo que el músculo sea estirado sin activar el reflejo al estiramiento.
Brodmann, K. (1905) Beiträge zur histologischen lokalisation der Grosshirnrinde. III. Die rindenfelder der niederen affen. J. Psychol. Neurol. 4: 177–226.
Brodmann, K. (1909) Vergleichende Lokalisationslehre der Großhirnrinde in ihren Prinzipien dargestellt auf Grund des Zellenbaues (Barth, Leipzig); English translation: Garey, L.J. (1994) Brodmann’s Localization in the Cerebral Cortex (Smith Gordon, London).
Brodmann, K. (1914) In Allgemeine Chirurgie der Gehirnkrankheiten (Eds. Knoblauch, A., Brodmann, K. & Hauptmann, A.) Pp. 86–426 (Verlag von Ferdinand Enke, Stuttgart).
Picard, N., and Strick, P. L. (1996). Motor areas of the medial wall: a review of their location and functional activation. Cereb. Cortex 6, 342–353.
Picard, N., and Strick, P. L. (2001). Imaging the premotor areas. Curr. Opin. Neurobiol. 11, 663–672.