Según el distinguido neurofisiólogo Rodolfo Llinás, en respuesta a las condiciones del medio ambiente un organismo como el nuestro sólo puede hacer dos cosas: salivar y moverse. Por supuesto, esta es sólo una forma jocosa de describir las funciones de un organismo, pero la realidad es que todas las actividades finales de un organismo son solamente de dos tipos: 1) la eliminación de productos intra- o extracelulares por secresión, excresión, varios tipos de transporte, etc., y; 2) movimientos internos y externos de traslación en el espacio.
Para llevar a cabo tanto las funciones necesarias para nuestra relación con el medio ambiente, todos los organismos animales con cierto nivel de complejidad han evolucionado un grupo de células que con sus prolongaciones forman redes, llamado sistema nervioso. Estas células, las neuronas, son diferentes de las otras células del cuerpo en que no solamente son capaces de generar y procesar información, sino también de pasar estas características a sus descendientes. Algunas de estas células se especializan en recoger información del medio ambiente, otras en procesarla para extraer de ella las porciones relevantes y otras mas en dirigir las actividades necesarias para responder a esa información en forma adecuada para mantener la vida del organismo y de la especie. Mas aún, como los organismos mas complejos están formados por numerosos órganos especializados en alguna función, el sistema nervioso también tiene que armonizar esas diferentes partes y mantener el organismo lo mas saludable posible, al menos hasta que cumpla su función de reproducción. Así, aunque a lo largo de la evolución desde los animales mas simples hasta los mas complejos, el sistema nervioso también ha sufrido enormes cambios, siempre ha tenido el mismo papel de relacionarlos con el medio ambiente y mantenerlos en condiciones óptimas para la vida y la reproducción.
Entonces, en esta sección veremos como el sistema nervioso origina y regula una de las dos actividades fundamentales de los organismos, la de los movimientos que producen desplazamientos en el espacio y que se deben a contracciones de músculos que llamamos esqueléticos porque están insertados en los huesos. Estos músculos también han sido llamados 'voluntarios', pero realmente no lo son, ya que el sistema nervioso no regula sus contracciones individuales, sino las de grupos musculares,que resultan en el desplazamientos de grandes regiones corporales, como el tronco, la cabeza, las extremidades y los dedos tanto de manos como de pies.
La iniciación del movimiento voluntario es un problema tanto fisiológico como filosófico. Esto es, cuando decimos que queremos iniciar un movimiento esto significa que al menos esa parte de los movimientos del organismo estará bajo el control de la voluntad, mientras que los que no lo están se denominan reflejos. Esta diferencia, movimiento voluntario versus movimiento reflejo, ha sido discutida durante siglos porque tiene como base el llamado 'libre albedrío', que es uno de los puntos principales de las religiones. O sea, si todo el comportamiento humano fuera simplemente el resultado de una cadena de reflejos, como fue propuesto inicialmente por Sechenov (1878), entonces el libre albedrío no existe y quedamos muy cerca de negar la responsabilidad de nuestras acciones.
Quizá el caso mas reciente relacionado con el problema del libre albedrío ocurrió en 1979 durante la llamada "Twinkie Defense", un caso en el que Daniel James White mató a George Moscone a Harvey Milk, el alcalde y el supervisor respectivamente de la ciudad de San Francisco, California, Estados Unidos. En su defensa los abogados arguyeron que White sufría de depresión concomitante con un cambio de dieta (de una saludable a otra formada con panecillos dulces, Twinkies), por lo que se encontraba en un estado como de 'zombie' y no era responsable de sus actos. Como resultado, White obtuvo una sentencia mucho menor a la esperada por un asesinato premeditado.
La historia de la discusión sobre el 'libre albedrío' es fascinante y, por supuesto, se inicia en el tan comentado trabajo de René Descartes (1596-1650), quien propuso que la estimulación sensorial es trasmitida a la glándula pineal, que selecciona y abre los 'tubos' nerviosos para llevar los espíritus animales hasta los músculos y producir su contracción. Años después Thomas Willis (1621-1675) colocó la conexión entre el alma y el cuerpo en el cuerpo calloso, proponiendo que la intensidad de las impresiones llega al estriado para determinar la ruta de los espíritus animales.
La discusión sobre el movimiento voluntario continuó y después de una serie de experimentos por Procháska, Whytt, Flourens, Goltz y otros, se llegó a la conclusión de que esos movimientos se asocian con movimientos espontáneos que no requieren estímulos sensoriales obvios, aunque no quedó claro dónde se originan. Sin embargo, ahora sabemos que indudablemente el origen de los comandos nerviosos que activan los músculos se encuentra en el SNC, por lo que podemos dividirlos en varias categorías:
1. Postura y locomoción. En muy pocas ocasiones los movimientos se originan en forma espontánea en animales crónicamente descerebrados. En cambio, la médula espinal y el tallo cerebral tienden a operar como mecanismos que en forma semejante a una máquina, inician movimientos que podemos llamar involuntarios.
2. Impulsos homeostáticos. Estos incluyen las desviaciones en los puntos de referencia de algunas variables internas, como la hipercapnia, hiperglucemia y desbalance electrolítico, que son poderosos motivadores para comportamientos como la respiración, alimentación y locomoción. Esas desviaciones son detectadas en el hipotálamo y otras áreas del tallo cerebral que conectan con el circuito motor. Así, como los movimientos que resultan tienen origenes detectables y generalmente son hechos en forma automática, caen en la categoría de movimientos involuntarios.
3. El tacto activo. Una gran parte de los movimientos de los animales son exploratorios y aunque frecuentemente es difícil detectar qué estímulos específicos los inician, estos movimientos voluntarios pueden verse mas bien como reflejos.
4. Atención, conciencia. Un atributo importante de los movimientos voluntarios es el que son concientes. Por ello, Crick (1984, 1995) sugirió que en el cerebro existe un foco de atención que ilumina procesamientos cognitivos particulares, como el 'libre albedrío', localizado en la parte anterior de la circunvolución callosa. Esto sugiere que existe un centro de control en el tálamo, ganglios basales y circunvolución callosa anterior, que 'atiende' a los eventos, definido esto como el proceso de selección y la capacidad de 'informar' sobre el blanco. Por ejemplo, las personas que sufren del 'síndrome de la Tourette' pueden describir sus movimientos involuntarios, por lo que la apreciación conciente del movimiento no parece necesaria para la volición. (Rothwell, en Prochazka et al., 2000)
Algunos fisiólogos dicen que realmente no existe un problema en todo esto, ya que las actividades estímulo/respuesta que hacen los sujetos siguiendo instrucciones son voluntarias, mientras aquellas respuestas que los sujetos no pueden modificar son reflejos. Sin embargo, la mayor parte de los movimientos que hacemos en la vida diaria son ejecutados sin que pongamos atención a ellos. Esto es, un movimiento como el levantar un brazo para agarrar un vaso con agua, está precedido y acompañado de una actividad fásica postural de los músculos de la espalda y a menos que se nos pregunte, no tenemos conciencia de esa actividad y no la podemos eliminar sin eliminar también los movimientos del brazo.
Entonces, definiremos como 'movimiento voluntario' aquel que es dirigido a un objetivo y que está completamente bajo control conciente, de manera que puede suprimirse voluntariamente, aunque el problema con esta definición es que muy pocos de nuestros movimientos ocurren en esta forma.
Esto nos lleva al tan discutido problema de qué es lo que se inicia en el sistema nervioso, por lo que tendriamos que mencionar aquellas regiones del SNC que son las primeras en activarse. Esto es, ¿cuáles son las regiones cerebrales que inician la cadena de actividades electroquímicas que en fisiología llamamos potenciales de acción y cómo viajan estas manifestaciones de actividad nerviosa a lo largo de los axones para llegar a los músculos e inducir su contracción?
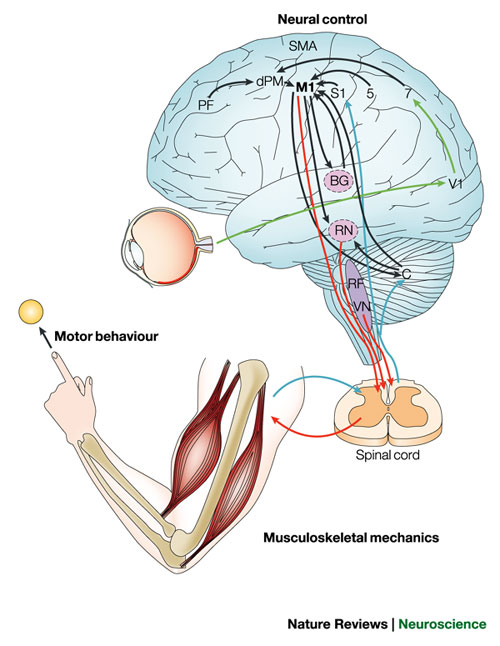
Las instrucciones para producir los movimientos voluntarios pasan por una etapa final en la corteza motora primaria, a la que llegan dos entradas importantes: 1) la corteza premotora, cuando los animales se mueven en respuesta a señales visuales y; 2) el área motora presuplementaria, cuando los animales hacen los mismos movimientos en forma espontánea, sin un estímulo inicial. Esta área suplementaria es el origen del llamado "potencial de disponibilidad", una descarga de actividad eléctrica en el cerebro durante el periodo justo antes de un movimiento voluntario.
Figura 1. Arriba. La corteza premotora prepara las instrucciones para la actividad voluntaria iniciada en respuesta a estímulos externos, mientras el área motora pre-suplementaria prepara las instrucciones para actividades 'intencionales' generadas internamente; ambas instrucciones son ejecutadas por la corteza motora primaria. Las señales que contienen copias de las instrucciones motoras preparadas también son mandadas a la corteza parietal, donde son usadas para predecir las consecuencias sensoriales del movimiento. Abajo. La preparación de las instrucciones motoras para los movimientos voluntarios por el área motora presuplementaria producen un sentido de urgencia. La parte inferior de la corteza parietal posterior genera representaciones sensoriales de las consecuencias predichas por el movimiento. (Haggard, 2009)
Es notable que cuando un cirujano estimula con un electrodo el área motora pre-suplementaria, el paciente indica que tiene "urgencia de moverse" (Fried et al., 1991), mientras cuando lo hace en la parte inferior de la corteza parietal posterior, el paciente también indica la experiencia de intención. Tradicionalmente la corteza parietal ha sido considerada un área de asociación sensorimotora que une estímulos visuales a las respuestas apropiadas, y es diferente de las áreas del lóbulo frontal responsables de la actividad voluntaria.
Hay diferencias importantes entre la estimulación frontal y la parietal, ya que la estimulación del área motora pre-suplementaria con corriente eléctrica poco intensa produce una experiencia de urgencia, mientras una estimulación mas fuerte produce movimientos (Fried et al., 1991). Esto sugiere que puede haber dos aspectos de la intención conciente (Figura 1), uno es la correlación conciente de preparar comandos motores en el área motora pre-suplementaria, mientras el otro es la predicción sensorial de las consecuencias de esos comandos en la corteza parietal.
Otras evidencias (Marsden et al., 1996; Rizzolati et al., 1996; Picard and Strick, 1996) sugieren la existencia de áreas mnotoras de órden superior, como el área motora suplementaria anterior (SMAa) y el área motora de la circunvolución callosa anterior (CMAa), que contribuyen con la mayor proporción de las fibras corticoespinales y están separadas de las áreas motoras corticales relacionadas con la ejecución del movimiento. Otras áreas, como el área motora primaria (MI), el área motora suplementaria posterior (SMAp) y el área callosa posterior CMAp, son llamadas áreas ejecutivas. Sin embargo, como también parecen participar en aspectos de órden superior del control motor, como la coordinación bimanual, parece que la distinción entre áreas motoras de órden superior y ejecutivas no es absoluta.
En la CMA se puede distinguir una parte anterior, una intermedia y una posterior, donde la anterior e intermedia son activadas en forma preferencial por tareas motoras que demandan la selección de parámetros de movimiento de naturaleza compleja, correspondiendo con su función de órden superior en el control motor. La parte posterior de la CMA puede ser activada durante movimientos simples iniciados externamente (Fink et al., 1997) y su arquitectura celular y conectividad son típicas de un área motora de salida. La CMA une el sistema límbico con las áreas motoras y mantiene la atención para actividades posteriores, por lo que sirve como una interface motivacional-motora.