En 1964 dos grupos de investigadores describieron potenciales corticales relacionados con los movimientos, uno de ellos es el 'Bereitschaftspotential' (Kornhuber y Deecke, 1964) y el otro es la 'variación contingente negativa' (Walter et al., 1964). Ambos potenciales reflejan cambios dinámicos en la actividad cortical unos 1.0-1.5 seg antes del movimiento, pero mientras el Bereitschaftspotential parece autónomo, la variación contingente negativa está relacionada con movimientos dirigidos. Estos dos trabajos estimularon una línea nueva de estudio de la fisiología motora hmana.
Aunque la primera descripción de la actividad electroencefalográfica que precede al movimiento voluntario en los humanos fue hecha por Kornhuber y Deecke (1964), ya antes Bates (1951) había intentado registrar la actividad relacionada con el movimiento por medio de una superposición fotográfica de numeros trazos del EEG, pero sólo pudo identificar la actividad que se produce después del movimento. Sin embargo, Kornhuber y Deecke registraron simultáneamente el EEG y el EMG mientras los sujetos repetían los movimientos a un paso lento y promediando el segmento del EEG previo al inicio del registro en el EMG, identificaron dos componentes del movimiento, uno antes y otro después del inicio del EMG. Estos fueron el 'Bereitschafspotential' (BP) o potencial de preparación (PP); Kornhuber y Deecke, 1964, 1965) y posteriormente los mismos autores encontraron dos componentes mas justo antes del inicio del movimiento, una positividad pre-movimiento (PMP) y un potencial motor (MP; Deeke et al., 1969).
Desde entonces, varios estudios sobre los potenciales corticales relacionados con el movimiento han mostrado esos potenciales, pero su significado fisiológico no es completamente claro todavía. En 1983 Libet y colaboradores usaron una nueva técnica en la que se pidió a los sujetos mirar un reloj y recordar el tiempo al cual estuvieron concientes de moverse e indicaron que esa intención ocurría mucho mas tarde que el BP. Ese trabajo cuestionó las implicaciones fisiológicas del BP, por lo que mas adelante veremos el problema con cierto detalle.
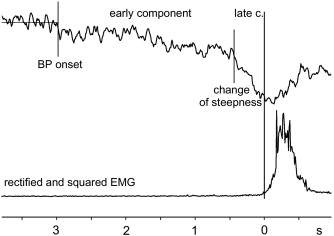
Se han propuesto varias terminologías para identificar los componentes de los potenciales corticales relacionados con el movimiento (MRCP), y uno de ellos que tiene 8 componentes (4 antes y 4 después del movimiento; Shibasaki et al., 1980) es utilizado en la Figura 1. En esta terminología cada componente, excepto BP y NS', dependen de la polaridad de la superficie y el intervalo promedio (en mseg) entre el pico de cada componente y el pico del EMG promediado. Este intervalo es negativo si el pico ocurre antes del pico del EMG y positivo si ocurre después del pico del EMG. Obviamente, el tiempo al pico de cada componente con respecto al inicio del movimiento depende de cómo se define ese inicio. En el caso de movimientos del dedo, el inicio del mecanograma es alrededor del mismo tiempo que el pico del EMG, pero está retrasado respecto al inicio del EMG por alrededor de 30 mseg. El BP se inicia alrededor de 2 mseg antes del inicio del movimiento y es máximo en el área centro-parietal y distribuído en forma simétrica independientemente del sitio del movimiento (Figura 2).
Figura 2. Potencial de preparación o "bereitschaftspotential". El trazo superior es el registro de la actividad cerebral (EEG), y el inferior es del músculo de la muñeca (EMG). El momento 0 es aquel en que se inicia el movimiento de la muñeca del sujeto experimental. Nótese que el registro EEG precede al momento del movimiento de la muñeca (tiempo 0) por casi 3 segundos, durante los cuales hay cambios eléctricos que pueden dividirse en componentes tempranos y tardíos. (Kornhuber y Deecke, 1964).
Entre los factores que influyen sobre la magnitud y curso temporal del BP registrado en condiciones de autofrecuencia están, el nivel de intención, el estado preparatorio, la selección del movimiento (seleccionado versus libre), el aprendizaje y la habilidad de la adquisición, la frecuencia de repetición, el esfuerzo percibido, la fuerza ejercida, la velocidad y precisión del movimiento, la terminación del movimiento y posibles lesiones en las estructuras cerebrales.
Se han usado varias técnicas para calcular las fuentes que generan el MRCP (Deecke y Kornhuber, 2003). En el caso de movimientos de la mano se calculó que tanto el SMA como la circunvolución precentral lateral, bilateralmente, eran las fuentes generadoras del BP temprano. Por otro lado, Praamstra et al (1996) estimaron tres fuentes bipolares para explicar el BP temprano, una en la SMA y las otras en la M1 bilateral. Mas aún, ellos determinaron que sólo la fuente de corriente identificada en el SMA era afectada por la selección del movimiento; esto es, era mayor antes del movimiento seleccionado libremente que con el movimiento fijo.
El Bereitschaftspotential podría ser originado por mas una sola fuente generadora que parece depender del tipo de movimiento. Por ejemplo, en el caso de un movimiento de las manos, Praamstra y colaboradores (1996) consideraron que la parte temprana del registro se debía a la actividad de tres fuentes, una en la SMA y las otras dos en la M1 bilateral. En cambio, Deecke y Kornhuber (2003) las encontraron bilateralmente en la SMA y la circunvolución precentral lateral, aunque sólo la fuente en la SMA era afectada por el tipo de movimiento y principalmente antes de un movimiento seleccionado libremente que de uno fijo. Por otro lado y con base en un análisis DC-EEG de alta resolución, Cui y Deecke (1999) demostraron que el BP ocurre inicialmente en las áreas motoras de la pared media (SMA y áreas mnotoras de la circunvolución del cíngulo) y después en la M1 contralateral, para localizarse finalmente en la M1 ipsilateral. Finalmente, Toma y colaboradores (2002) han usado el análisis de componentes principales y la resonancia magnética funcional para determinar que la fuente principal de la parte inicial del BP es la parte superior de la circunvolución precentral bilateral, específicamente, el área de la mano en el área 6 y el área 4, como la fuente de la porción tardía del registro y el área 3 como la fuente del fpMP o N+50. Esto es, en la mayor parte de estos estudios la fuente de MP o de N-10 ha sido localizada en el área de la mano de M1.
Con base en los resultados descritos arriba, se ha postulado que el BP temprano refleja la preparación subconciente para ejecutar el movimiento, mientras que el BP tardío refleja el deseo conciente de efectuar ese movimiento.

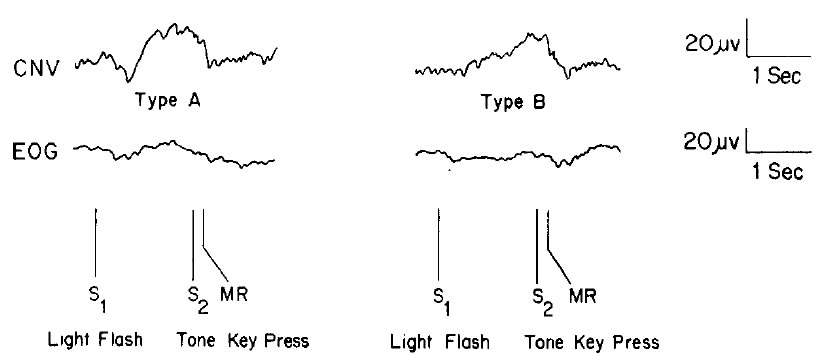
Figura 4. Registro poligráfico de dos tipos de VCN basados en un tiempo de subida rápido (tipo A) y lento (tipo B). El trazo inferior es el electrooculograma (EOG). S1 indica el momento del destello de luz, S2 el tono que indica apretar la llave, y MR el movimiento de la mano. Notese que la VCN se encuentra entre S1 y S2. (Figura 3; Tecce, 1972)
Bates, J.A.V. 1951. Electrical activity of the cortex accompaning movement. J. Physiol. 113: 240-257.
Brodmann, K. 1909. Vergleichende Lokalisationslehre der Grosshirnrinde in ihren Prinzipien dargestellt auf Grund des Zellenbaues. Leipzig: Barth.
Cui, R.Q., and L. Deecke. 1999. High resolution DC-EEG analysis of the Bereitschaftspotential and post movement-related potentials to illuminate human information processing. Biol. Psychol. 26: 69-89.
Deecke, L., P. Scheid, and H.H. Kornhuber. 1969. Distribution of readiness potential, pre-motion positivity and motor potential of the human cerebral cortex preceding voluntary finger movement. Exp. Brain Res. 7: 158-168.
Deecke, L., and H.H. Kornhuber. 2003. Human freedom, reasoned will, and the brain: The Bereitschaftspotental story. In: Jahanshashi, M., and M. Hallett, editors. The Bereitschaftspotential. Movement-related cortical potentials. New York: Kluver Academic/Plenum Publishers; p. 283-320.
Donchin, E. 1979. Event-related bain potentials: a tool in the study of human information processing. In: Evoked potentials and behavior. (Ed. H. Begleiter), pp. 13-75. Plenum, New York.
Kornhuber, H.H., and L. Deecke. 1964. Hirnpotentialänderungen beim Menschen vor und nach Willkurbewegungen, dargestellt mit Magnetband-Speicherung und Ruckwartsanalyse. Pflügers Arch. 281: 52.
Kornhuber, H.H., and L. Deecke. 1965. Hirnpotentialänderungen bei Wilkurbewegungen und passiven Bewegungen des Menschen: Bereitschaftspotential und reafferente Potentiale. Pflügers Arch. 284: 1-17.
Libet, B., C.A., Gleason, E.W. Wright, and D.K. Pearl. 1983. Time of conscious intention to act in relation to onset of cerebral activity (readines-potential). The unconscious initiation of a freely vountary act. Brain 106: 623-642.
Pramstra, P., D.F. Stegeman, M.W. Horstink, and A.R. Cools. 1996. Dipole source analysis suggests selective modulation of the supplementary motor area contribution to the readiness potential. Electroencepahalogr. Clin. Neurophysiol. 98: 468-477.
Shibasaki, H., G. Barrett, E. Halliday, and A.M. Halliday. 1980. Components of the movement-related cortical potential and their scalp topography. Electroenceph. Clin. Neurophysiol. 49: 213–26.
Shibasaki, H., and M. Hallett. 2006. What is the Bereitschaftspotential? Clin. Neurophysiol. 117: 2341-2356.
Tecce, J.J. 1972. Contingent negative variation (CNV) and psychological processes in man. Psychol. Bull. 77(2): 73-108, 1972.
Toma, K., M. Honda, T. Hanakawa, T. Okada, H. Fukuyama, A. Ikeda, et al. Activities of the primary and supplementary motor areas increase in preparation and execution of voluntary muscle relaxation: an event-related fMRI study. J. Neurosci. 19: 3527-3534.
Walter, W.G., R. Cooper, V.J. Aldridge, W.C. McCallum, and A.L. Winter. 1964. Contingent negative variation: an electric sign of sensorimotor association and expectancy in the human brain. Nature 203(4943): 380-384.