
El soma de una célula nerviosa típica tiene dimensiones microscópicas (en promedio entre 10 y 30 µm de diámetro) pero el poder de la batería generada a través de su membrana externa es sorprendentemente grande, unos 100 mV, lo cual es poco menos que un décimo de una pila ordinaria. Esa potencia en un elemento tan pequeño genera la pregunta, ¿de dónde sale la energía para hacerla?
Para responder esa pregunta debemos buscar dentro o fuera de la célula 'cosas' que estén relacionadas con cargas eléctricas, ya que para producir una señal eléctrica tiene que haber una corriente eléctrica, que no es otra cosa sino el flujo de partículas cargadas, como electrones o iones. Como los electrones no se encuentran libres en las células biológicas, debemos buscar iones adentro y afuera, y varios de ellos pueden contribuir al establecimiento de la batería eléctrica, entre ellos los iones de potasio, sodio, calcio y cloro, y cuando medimos esos iones tanto en el interior como en el exterior de la célula, podemos hacer varias observaciones interesantes:
1) En el exterior de la célula la concentración de iones positivos (cationes) es igual a la de iones negativos (aniones), por lo que hay electroneutralidad en el líquido extracelular.
2) Adentro de la célula el número de cationes y aniones es igual, lo que quiere decir que también hay electroneutralidad.
3) La concentración de iones Na+ en el exterior de la célula es mucho mayor que en el interior.
4) La concentración de iones K+ adentro de la célula es mucho mayor que adentro.
5) La concentración de iones Cl- fuera de la célula es mayor que en el interior.
Figura 1. Diferencias en las concentraciones iónicas entre el interior (amarillo) y el exterior (azul) de una célula. Notese que en el interior predominan los iones K+ y A-, mientras en el exterior los iones mas comunes son Na+ y Cl-. Sin embargo, en ambos compartimentos hay electroneutralidad.
Las observaciones anteriores nos dicen que la concentración de iones no es igual adentro y afuera de la célula, lo que nos explicamos por la presencia de la membrana, ya que si esta no existiera los iones estarían libres para penetrar al interior de la célula e igualarían sus concentraciones (claro que si no hubiera membrana tampoco habría célula).
El gradiente iónico a través de la membrana es una forma de energía y es equivalente a tener un tanque lleno de agua a cierta altura sobre el suelo, de manera que al abrir la llave el agua empezará a salir con una fuerza que dependerá de la altura a la que se encuentre el tanque. Sin embargo, a diferencia de las moléculas de agua, los iones son partículas cargadas, por lo que en la célula tenemos dos fuerzas, una fuerza química debida al gradiente de concentración y una fuerza eléctrica debida a la carga de los iones. Cuando los iones se mueven empujados por su gradiente de concentración también llevan su carga, de manera que, por ejemplo, el movimiento de iones Na+ lleva cargas positivas a la región donde llegan.
Para entender claramente el comportamiento de los iones dentro y fuera de la célula podemos hacer un experimento simple, que involucra un vaso con agua, un huevo y un voltímetro para medir el potencial. Primero hacemos pequeños agujeros en los dos extremos del cascarón de un huevo de gallina, dejando intacta la membrana en el compartimento con aire y abriéndola en el otro extremo. Después de vaciar y lavar el huevo, lo colocamos en un soporte dentro de un vaso y simultáneamente ponemos en el compartimento exterior (e) una sal simple (KCl) a una concentración de 100 mM (o 100 partículas de cada uno si se prefiere) y en el compartimento interior (i) una sal diferente, hecha con un poco de proteina (P-) y Na+ a una concentración también de 100 mM (o si se prefiere, 100 particulas de K+ y 100 de P-). Diremos que la membrana que separa los dos lados es semipermeable, esto es, que permite a los iones K+ y Cl- pasar a través de ella, pero no a las P- porque son moléculas muy grandes.

Figura 2. Difusión de iones positivos a través de la membrana cuando hay canales permeables a ellos. Su movimiento está impulsado por el gradiente de concentración a ambos lados de la membrana. Para simplificar, en la Figura no se ven los iones negativos, pero suponemos que hay electroneutralidad a cada lado de la membrana.
Notaremos que el número de cargas positivas y negativas a cada lado de la membrana es la misma (100 y 100), por lo que habrá electroneutralidad tanto en el compartimento interior como en el exterior. Notaremos también que no hay diferencia en las concentraciones de iones positivos a través de la membrana.
Ahora, lo que ocurrirá después de esas condiciones iniciales es lo siguiente.
1. Cuando no hay barreras a la difusión, las moléculas se mueven de la región donde están más concentradas a donde lo están menos y como en este caso los iones Cl- encuentran en la membrana canales que les permiten el paso, entrarán al huevo llevando con ellos su carga negativa. Como resultado de ese movimiento habrá más cargas negativas en el interior que en el exterior del huevo, por lo que el número de cargas positivas en el exterior será mayor que en el interior. Así, el compartimento interior se hará negativo con respecto al exterior.
2. Conforme los iones Cl- se mueven hacia el interior del huevo generan un gradiente eléctrico que hace el interior sea negativo respecto al exterior y esa negatividad atrae los iones K+ del exterior.
3. Los iones K+ empezarán a moverse hacia el interior del huevo y al hacerlo, neutralizarán algunas de las cargas negativas del Cl-, pero al mismo tiempo aumentarán la concentración de K+ en el interior, comparada con la del exterior. Entonces, conforme la negatividad del interior empieza a ser neutralizada por los iones K+ que entran, los iones Cl-, que habían dejado de entrar repelidos por la negatividad del interior, empezarán a moverse nuevamente hacia dentro del huevo. Esto atraerá más iones K+ y así sucesivamente, hasta que se establezca un estado de equilibrio, en el que ya no habrá movimiento neto de ninguno de los iones K+ y Cl-.
Por lo tanto, en el estado de equilibrio tenemos las siguientes condiciones:
a) Habrá más iones K+ en el interior del huevo que en el exterior.
b) El interior del huevo será ligeramente mas negativo que el exterior.
c) La mayor parte de los iones Cl- permanecerán en el exterior y habrá una concentración ligeramente menor en el interior.
d) No habrá cambios en la concentración de las P- que permanecerán en el interior.
Las condiciones que hemos descrito para el experimento hecho con el huevo de gallina son extraordinariamente semejantes a las que se encuentran en las células de los tejidos biológicos. En ellas, la concentración de K+ es mayor en el espacio intracelular que en el extracelular y en este la concentración de Cl- es mayor que en el interior. Además, cuando se mide con microelectrodos intracelulares, el interior de la célula es más negativo que el exterior, por lo que el modelo del huevo nos ha dado una buena idea sobre como se establece el potencial transmembrana en una célula biológica. Las diferencias que aún se pueden ver entre las células de los tejidos biológicos y el huevo se deben principalmente a las concentraciones de iones y los valores del potencial, por lo que no son fundamentales.
Para describir la situación real en las concentraciones iónicas de las células de los tejidos, deberiamos tener otro elemento en la membrana, una maquinaria que evite que las concentraciones iónicas a ambos lados de la membrana varíen y por lo tanto mantenga el estado estacionario del potencial de reposo. Este mecanismo es llamado 'bomba' de Na+/K+, que toma los iones sodio que contínuamente entran a la célula y los saca, al mismo tiempo que mete los iones K+ que hayan salido. El resultado de esta acción de bombeo es que las concentraciones iónicas en ambos lados de la membrana son mantenidas es un estado estacionario durante tanto tiempo como viva la célula.

Figura 3. Diagrama de una 'bomba' iónica que mueve iones Na+ desde el interior de la célula al exterior y también iones K+ desde el exterior hacia el interior. Nótese que el número de iones movidos no es igual en ambas direcciones. La molécula roja (P) indica que para su funcionamiento la bomba requiere la energía del ATP.
Entonces, el potencial eléctrico a través de la membrana de las células vivas tiene las siguientes características:
1. Está relacionado con el gradiente de concentración a través de la membrana celular para los iones que pueden pasar por ella a través de canales específicos. Esto se debe a la permeabilidad selectiva de la membrana.
2. Es mantenido por la bomba Na+/K+ que funciona usando energía metabólica.
3. Cambia con los gradientes de concentración o los canales que permiten el paso de iones a través de la membrana.
4. Es un proceso que sigue las reglas de la difusión y las fuerzas electroquímicas.
El potencial de la membrana en las condiciones de equilibrio para cada ión también puede calcularse con la ec. de Nernst, que derivamos a continuación.
La ecuación de Nernst nos proporciona el voltaje al cual se habrá llegado a un potencial de membrana, llamado "potencial de equilibrio", al cual por cada ión que entra a la célula habrá uno que sale y por lo tanto el potencial no cambiará. Este concepto de tal importancia que en esta sección derivaremos la ecuación de Nernst, esperando que esto sirva para ayudar a entender plenamente el concepto de potencial de equilibrio.
Empezaremos por definir la diferencia de energía potencial eléctrica de 1 mol de K+, que es el trabajo requerido para mover 1 mol de K+ a través de la membrana, en contra de una diferencia de potencial eléctrico dado por V1-V2. Por ejemplo, en el caso de una célula esto sería el trabajo requerido para mover los iones K+ desde el exterior hasta el interior de la célula en contra del gradiente eléctrico, un trabajo dado por,
![]()
donde,
WE - trabajo
ZK - valencia del ion K (+1)
F - constante de Faraday (96,487 Coulombs/mol)
(V1-V2) - diferencia de potencial (Joules/Coulombs = Volts)
En forma semejante, el trabajo requerido para llevar 1 mol de K+ desde el exterior hasta el interior de una célula en contra del gradiente de concentración, es la diferencia de potencial químico debido a la distribución desigual de K+ y se describe con la siguiente ecuación.
![]()
Esto es, el trabajo hecho para mover los iones K+ en contra de sus gradientes eléctrico y de concentración es la suma de ambos trabajos y se describe como:
![]()
![]()
Ahora, cuando los iones K+ se encuentran en equilibrio a través de la membrana ΔWK = 0 y V1-V2 es el potencial transmembrana Vm. Despejando V men la ecuación (3) tenemos,
![]()
Esta es la ecuación de Nernst y proporciona el valor del potencial (Vm) al cual los iones K+ se encuentran en equilibrio a través de la membrana; esto es, cuando no existe flujo de potasio. Este potencial de membrana es también llamado el 'potencial de equilibrio del ion potasio (EK)'. Por esto, cuando un ion se encuentra a su potencial de equilibrio se dice que se encuentra distribuído pasivamente.
Otra derivación mas completa de la ecuación de Nernst, a partir de los flujos iónicos, puede encontrarse aquí
La hipótesis de que la ecuación de Nernst describe el potencial de la membrana dado por el potasio cuando la neurona está en reposo, fue validada midiendo el potencial de reposo del axón gigante del calamar por medio de electrodos en su interior, encontrándose que su valor (potencial transmembrana, Vm) era de -65 a -70 mV, cercano, pero no exactamente igual al potencial de equilibrio (EK+) calculado para los iones K+. Por lo tanto, Hodgkin y Horowics decidieron estudiar la influencia de la concentración externa de potasio sobre el potencial de reposo aumentando la concentración de [K+] y después de probar numerosas concentraciones, mostraron que los resultados caían en una línea con una pendiente de -58 mV (Figura 2). Este resultado demuestra que,
- cada vez que el potasio extracelular aumenta diez veces el potencial aumenta 58 mV.
- la línea queda muy cercana a los números predichos por la ecuación, excepto a concentraciones externas bajas de K+.
- hay otros iones que influyen sobre el potencial de reposo de la membrana.
- la membrana también es permeable a los iones Na+ y al Cl-, pero este último tiene poca influencia sobre el potencial de membrana.
- la permeabilidad al Na+ es mucho mas baja que a los iones K+.
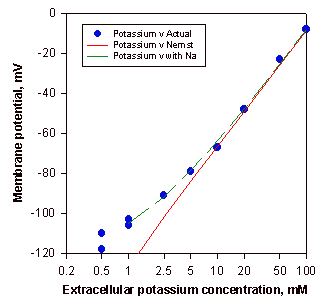
Figura 2. Resultado de cambiar la concentración extracelular de K+ sobre el potencial de membrana. Los puntos son mediciones experimentales del potencial de reposo del axón gigante del calamar al cambiar las concentraciones externas de K+. La línea recta es la solución de la Ec. de Nernst para el potencial de equilibrio del K+ a esas concentraciones de K+ y predice los resultados a concentraciones extracelulares de K+ moderadas y altas. La desviación a concentraciones bajas de K+ se debe a que este deja de ser el ión predominante de la solución externa y empieza a verse la influencia de otros iones, como el Na+ y el Ca++ (Hodgkin y Horowicz).
La permeabilidad se debe a la presencia de canales que permiten que los iones entren o salgan de la célula, de manera que,
1. Los iones Na+ entran a la célula debido a su gradiente de concentración.
2. Los iones Na+ producen una ligera depolarización de la membrana, alejándola del potencial de equilibrio del K+.
3. Por lo tanto, el Vm del axón gigante del calamar es de -65 mV en vez de -93 mV.
4. En el equilibrio las corrientes llevadas por la entrada de Na+ y la salida de K+ están balanceadas y el potencial de reposo es estable.