La membrana de las neuronas se comporta como un circuito eléctrico simple que tiene una resistencia y un capacitor en paralelo, y para entender su comportamiento es útil revisar el comportamiento de algunos cicuitos eléctricos relacionados. Sin embargo, primero veremos algunos conceptos y definiciones.
Cuando dos cargas diferentes son separadas en el espacio se produce un voltaje o diferencia de potencial y entre mayor sea la cantidad de carga separada, mayor será el voltaje V y mayor la tendencia de las cargas a volverse a reunir. El voltaje siempre se mide en un punto con respecto a otro y no puede haber voltaje en un solo punto en el espacio. La unidad de voltaje es el Volt.
El flujo de cargas eléctricas es la corriente, que se mide en Coulombs/segundo o Amperes. La resistencia, R, es una medida de la dificultad de la corriente para fluir en un circuito y entre mayor sea esta dificultad, mayor será la resistencia. La unidad de resistencia es el Ohm, Ω. El recíproco de la resistencia es la conductancia, g, una medida de la facilidad con la que la corriente fluye en un circuito. La unidad de conductancia es el Siemen (el término antiguo era mho).
La corriente, el voltaje y la resistencia, están relacionados uno con otro por la ley de Ohm (I = V/R). Esta relación para circuitos resistivos simples dice que, para una resistencia dada, la corriente aumenta linealmente con el voltaje o, alternativamente, la corriente producida por un voltaje particular depende de la resistencia a ella. Para los que no están familiarizados con conceptos eléctricos, una analogía útil podría ser el llevar piedras mientras subimos una colina. La corriente puede ser el número de piedras que llevamos por viaje, la resistencia sería la pendiente de la colina y el voltaje la cantidad de energía que usamos para llevar las piedras. Es claro que entre menor sea la pendiente (menor la resistencia) y entre mas grande nuestro nivel de energía (mayor el voltaje), mayor será el número de piedras que podemos llevar hasta la parte superior de la colina. Por supuesto, la ley de Ohm puede ser reescrita como I = gV, donde g = 1/R. En esta forma de la ecuación, la corriente es una función directa del voltaje y de la conductancia.
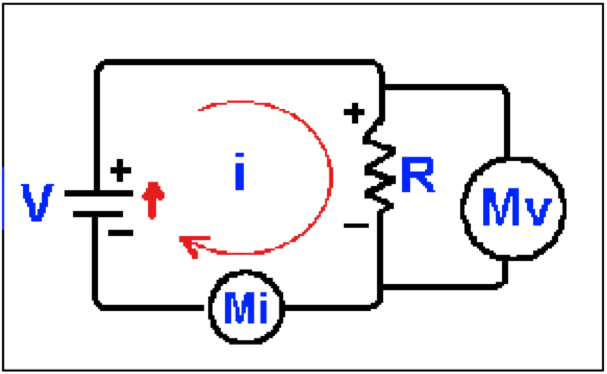
Figura 1. Un circuito eléctrico con una resistencia simple. Usamos una batería, V, para aplicar un voltaje de 10 V a ambos lados de una resistencia, R, de 10 Ω. Medimos el voltaje con un voltímetro, Mv, en paralelo con R, y la corriente con un amperímetro, Mi, en serie con R. En este caso el voltímetro lee 10 V, con la polaridad indicada por los signos + y -, mientras el amperímetro lee 1 A.
Consideremos el circuito de la Figura 1, que contiene un elemento activo, la fuente de voltaje, V, indicada por la batería y un elemento pasivo, la resistencia R. Pensemos que la fuente de voltaje proporciona corriente o energía, mientras la resistencia es el consumidor de esa energía. La ley de Kirchoff dice que la corriente fluirá sólo en circuitos que están completos o cerrados y podría ser útil pensar en como fluye el agua por un tubo, ya que si el tubo no tiene hoyos, toda el agua que entra en un extremo deberá fluir hasta el otro extremo. Como tenemos una diferencia de potencial y una resistencia, sabemos que habrá corriente (i) en el circuito y que puede ser medida por el amperímetro, Mi.
Esa corriente puede ser calculada con la ley de Ohm:
Si V = 10 V
y R = 10 Ω
Entonces, ![]()
La corriente que fluye por la resistencia produce una caída de voltaje (una diferencia en el potencial eléctrico) a través del resistor, que puede ser medida con el medidor Mv , un voltímetro. Podemos suponer que el medidor es perfecto (esto es, que no hay corriente fluyendo a través de él) y que no tiene influencia sobre la operación del circuito, por lo que todo el voltaje V será impuesto a la resistencia R. La caída de voltaje medida a través del resistor R será 10 V, con la polaridad que se muestra en la Figura 1. Como los alambres del circuito tienen muy poca resistencia no habrá caída de voltaje debido a ellos (a partir de la ley de Ohm, V = IR y si R = 0, entonces V = 0) y el potencial será el mismo en los exremos superiores del resistor y la batería. El mismo argumento se aplica a los extremos inferiores.
Por convención, se dice que la dirección del flujo de corriente en el circuito externo, la parte del circuito fuera de la batería, es la dirección en la que se mueve una partícula positiva (flecha delgada semicircular). De hecho, en este tipo de circuitos la corriente es llevada por electrones, pero la dirección del flujo de corriente es opuesta a su movimiento; esto es, será de + a -. Para completar el circuito la corriente debe fluir de - a + dentro de la batería (el circuito interno). En contraste con esta corriente eléctrica, que es llevada por electrones, la mayor parte de los transportadores de carga en los eventos membranales son partículas cargadas positivamente o cationes y, por convención, las corrientes fluirán en la dirección en que se mueven los cationes. Hay que pensar que cuando un anión o un ión cargado negativamente se mueve, se dice que la corriente va en dirección opuesta a su movimiento y frecuentemente esto da problemas a los estudiantes cuando consideran los iones Cl-.
Notemos también que cuando la corriente entra al resistor (abajo en la Figura 1), ese extremo del resistor se hace positivo con respecto al otro extremo. Como veremos después, la corriente que sale a través de la membrana en reposo hará la membrana mas positiva en el interior respecto al exterior y lo inverso será verdad para corrientes que se dirigen hacia dentro de la célula.
Esta observación es verdad para flujos de corriente en todos los elementos pasivos, como resistores y membranas celulares en "reposo". En relación a la Figura 1, vemos que la corriente en la fuente o elemento activo (la batería) va de negativo a positivo (la flecha pequeña, gruesa). Por lo tanto, una corriente de entrada a través de una membrana activa hará la membrana mas positiva en el interior con respecto al exterior; esto es, la hipopolarizará. Veremos que una corriente de sodio entrante hipopolariza la membrana del nervio activo, mientras una corriente saliente en la membrana en reposo (pasiva) la hiperpolariza. Esto puede resumirse diciendo que, "las corrientes salientes hipopolarizan membranas pasivas, mientras las corrientes entrantes hipopolarizan membranas activas."
Un capacitor es una estructura capaz de separar y almacenar carga y generalmente está hecho con dos placas conductoras paralelas separadas por un material no conductor o dieléctrico. La capacitancia de un capacitor es simbolizada con la letra C y está relacionada con el voltaje por la ecuación C = Q/V, donde Q es la carga y la unidad de capacitancia es el Faradio. Entre mas grande sea la capacitancia mayor será la cantidad de carga que debe ponerse en las placas para llevar el voltaje entre ellas a un valor dado. Visto en otra forma, entre mayor sea la capacitancia mayor será la cantidad de carga almacenada en las placas para un voltaje dado entre ellas. Nos referimos a aumentar la carga almacenada en un capacitor como "cargar el capacitor" y a disminuir la carga almacenada como "descargar el capacitor".
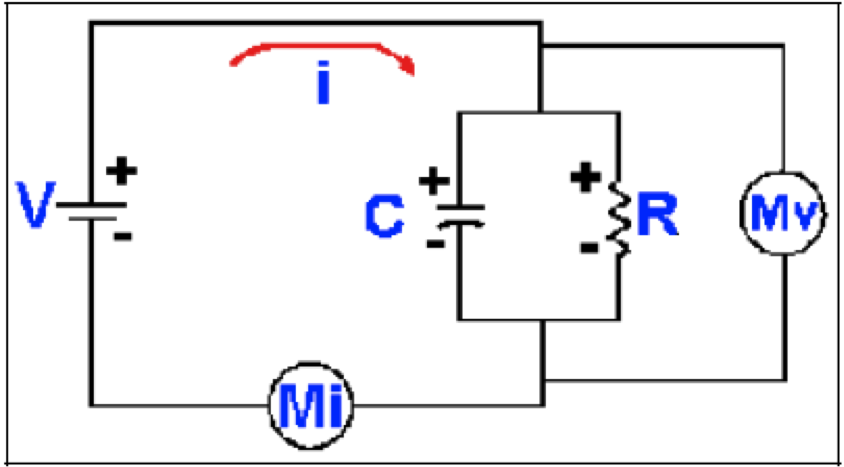
Figura 2. Un circuito simple con resistencia y capacitancia. Este es el mismo circuito de la Figura 1, pero con un capacitor, C, colocado en paralelo con la resistencia, R. El voltaje a través del resistor y el capacitor es el mismo voltaje que el de la batería, V, y el mismo que se lee en el voltímetro, Mv. La polaridad del voltaje está indicada por los signos + y -.
La introducción de un capacitor, como en la Figura 2, cambia el comportamiento del circuito. Como el capacitor tiene un dieléctrico entre las placas, no hay electrones fluyendo entre ellas, pero cuando se conecta la batería al circuito las cargas positivas que se acumulan en la placa superior son llevadas desde la placa inferior, lo que produce un voltaje entre ellas. La carga que fluye hacia o desde la placa es una corriente, la corriente capacitiva ic, que está relacionada a la capacitancia y voltaje por la ecuación:
ic = C dV/dt
Si no hubiera una resistencia en el circuito de la Figura 2 la batería podría ser desconectada y el voltaje entre las placas del capacitor permanecería en V (al menos para un capacitor perfecto). El resistor proporciona el camino para la corriente a través del cual se descargará el capacitor, con la corriente fluyendo desde la placa positiva a la placa negativa. Entre mayor sea la R menos corriente fluirá por unidad de tiempo y mas lentamente caerá el voltaje del capacitor. De hecho, el tiempo para descargar el capacitor varía directamente con R y C. El tiempo requerido para que el voltaje llegue a (1-1/e) ΔV o aproximadamente b ΔV (ΔV es el cambio total de voltaje) es llamado la constante de tiempo, τ, y es dado por la ecuación τ = R x C. Si R se expresa en Ohms y C en Faradios, entonces τ estará en segundos. En circuitos como el de la Figura 2, sin rectificación, las constantes de tiempo para cargarse y descargarse son iguales. Sin embargo, como veremos, las membranas celulares tienen propiedades rectificantes que hacen estos tiempos diferentes.
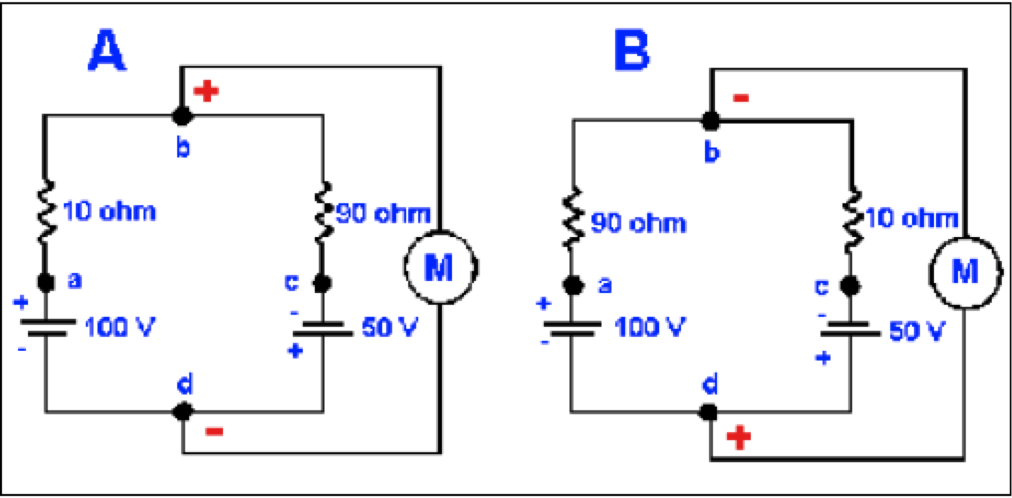
Figura 3. Un circuito simple con resistencias en paralelo. Las dos resistencias, R1 y R2 están en paralelo; esto es, comparten las mismas entradas y combinan sus salidas. El voltímetro, M, lee el valor de la batería V.
En los circuitos eléctricos las resistencias en serie simplemente se suman. Así, la R en la Figura 1 puede ser reemplazada por dos resistores de valor R/2 en serie (i.e., la salida de uno será la entrada del otro) sin cambiar las propiedades del circuito. Sin embargo, en las resistencias colocadas en paralelo y que comparten la misma entrada y combinan sus salidas, ya no puede hacerse eso.
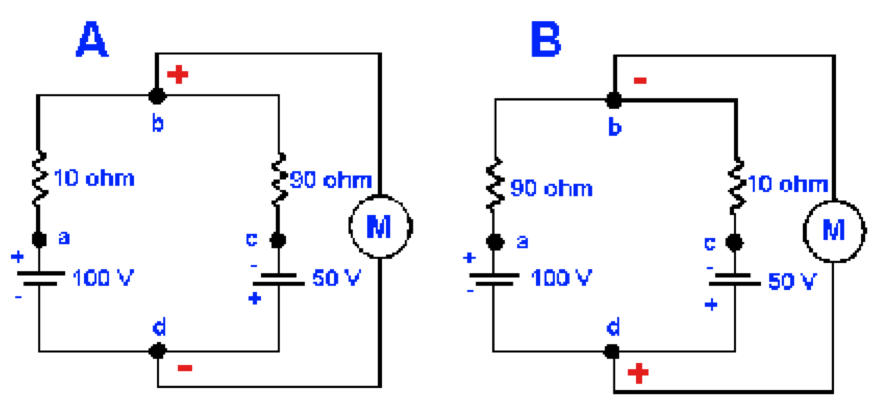
Figura 4. Efectos de variar la resistencia sobre el voltaje de un circuito. En cada circuito se muestra una batería y un resistor en paralelo. A. En esta configuración el voltímetro, M, leerá 85 mV positivo en el punto b respecto al punto d. B. El mismo circuito que en A, pero con las resistencias intercambiadas, lo que hace que el voltímetro lea 35 V, negativo en el punto b respecto al punto d. Esto es, simplemente cambiando los resistores la polaridad y el votaje han cambiado.
Consideremos el circuito resistivo paralelo de la Figura 4. La diferencia de potencial a través de R1 y R2 será la misma, V. Así, la corriente a través de R1 será
![]()
y la corriente a través de R2 será
![]()
![]()
y dividiendo entre V obtenemos
![]()
Por lo tanto, en un circuito resistivo paralelo las resistencias se suman reciprocamente. En cambio, cada uno de los capacitores en paralelo puede almacenar carga, por lo que para un mismo voltaje almacenarán mas carga. En paralelo Ctotal = C1 + C2, pero cuando los capacitores están en serie, cada uno de ellos ve un voltaje que es menor y por tanto almacenará menos carga. Cuando los capacitores están en serie:
![]()
Todavía debemos considerar dos circuitos mas antes de discutir los eventos membranales y que se muestran en la Figura 5. Nótese que en A las baterias mueven corriente en la misma dirección, por lo que el voltaje entre los puntos a y c será simplemente la suma de los voltajes de las dos baterías, 150 mV, con el punto a positivo respecto al punto c. La resistencia total del circuito (los resistores están en serie) es 100 Ω. Por lo tanto, la corriente en el circuito es 150 V/100 Ω o 1.5 A. Cuando 1.5 A fluyen a través de 10 Ω hay una caída de potencial de 1.5 A x 10 Ω y se producen 15 V, con el punto a positivo respecto al punto b. La diferencia de potencial entre los puntos b y cserá mostrado en el medidor, 100 V - 15 V = 85 V, con el punto b positivo respecto al punto c. Los cálculos de voltaje usando la porción de 90 Ω del resistor del circuito deben producir el mismo valor.
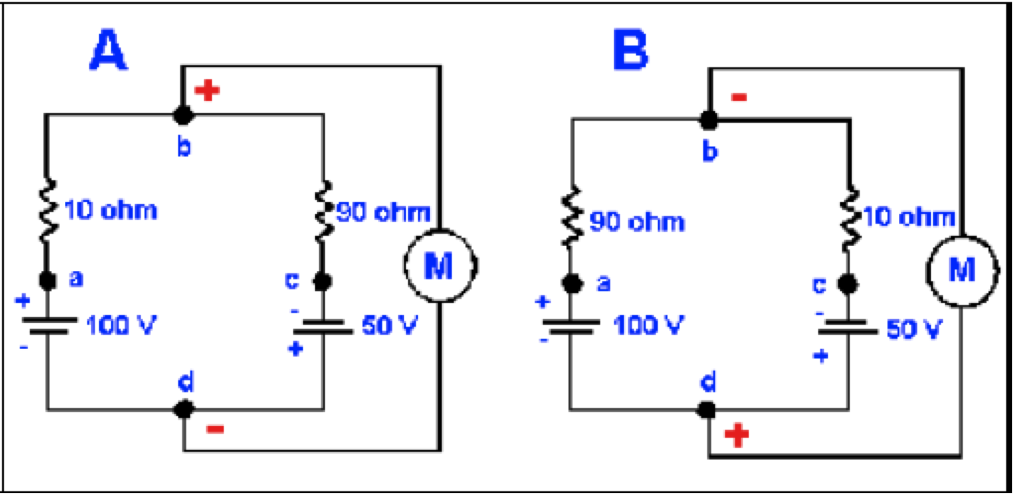
Figura 5. Efectos de variar la resistencia sobre el voltaje del circuito. Se muestra una batería y un resistor en cada circuito paralelo. A. Con esta configuración el voltímetro, M, leerá 85 V, con el punto b positivo respecto al punto d. B. El mismo circuito que en A pero con las resistencias intercambiadas, llevando al voltímetro a leer 35 V, con el punto b negativo respecto al punto d. La polaridad y el voltaje han cambiado simplemente cambiando las reistencias.
La Figura 5B es la misma que la Figura 5A, excepto que los valores de las resistencia están intercambiados. La corriente en el circuito es aún 1.5 A, pero esta vez el medidor leerá 50V - 15V = 35V, con el punto b negativo respecto al punto d. De nuevo, por simetría, el cálculo para la resistencia de 90 Ω dará el mismo valor. Nótese que la diferencia de potencial entre los puntos b y d cambia en magnitud y polaridad simplemente cambiando la resistencia (o conductancia), sin alterar los voltajes de las baterías. Esta observación es importante cuando se considera el orígen del potencial de acción.

Figura 6. Efectos de la resistencia y la capacitancia de la membrana sobre el voltaje transmembrana. A la izquierda se muestran dos microelectrodos penetrando un axón, uno para estimular (derecha) y otro para registrar (izquierda). El potencial transmembrana se registra entre el microelectrodo dentro del axón (izq) y otro electrodo fuera del axón (botón negro), usando el amplificador A y el resultado se ve en un osciloscopio, M. La corriente estimulante fluye desde el segundo microelectrodo, sale a través de la membrana y regresa a la batería por el otro electrodo externo. La distancia entre las micropipetas puede cambiar. En el trazo superior a la derecha se ve la corriente inyectada y en los otros trazos el voltaje transmembrana cuando los electrodos están separados por 0, 0.5, 1.0 y 2.0 mm respectivamente.
Eléctricamente el axón se comporta como un conductor de electricidad muy pobre, ya que el axoplasma tiene una gran resistencia. Por ejemplo, la resistividad longitudinal del axón gigante del calamar es de 30-60 Ωcm, lo que es extremadamente alto comparado con la de un alambre de cobre del mismo diámetro, que tiene una resistividad de 1.8 x 10-6 Ωcm. Esta diferencia en la resistencia se debe a que en el axón la densidad de cargas es pequeña (los iones en el axoplasma tienen menor densidad que los electrones en el alambre de cobre) y a que su movilidad es menor (los iones en solución se mueven mucho menos que los electrones en un alambre de cobre). Por lo tanto, la caída de voltaje en 1 cm de citoplasma es alrededor de 107 veces mayor que la que ocurre a lo largo de 1 cm de alambre.
La membrana de una neurona se comporta como una resistencia; esto es, cuando se pasa corriente a través de la membrana, hay una caída de voltaje que se predice con la ley de Ohm. Generalmente los valores de la resistencia de la membrana son expresados como una resistencia específica, en Ωcm2. (La resistencia de la membrana disminuye con el aumento en el área de membrana; por lo tanto, es medida en Ω x cm2, mientras la capacitancia de la membrana aumenta con el aumento en el área y por lo tanto es expresada en µF por cm2). Las motoneuronas de la langosta (una neurona grande en la que fácilmente se pueden hacer mediciones) tienen una resistencia de la membrana de 2,300 Ωcm2.
La membrana neuronal también se comporta como un capacitor, esto es, es capaz de separar y almacenar carga. La capacitancia de la mayor parte de las membranas neuronales es del orden de 1 µF/cm2, lo que quiere decir que la membrana puede separar y almacenar una carga de 1 x 106 coulombs/volt de potencial a través de cada cm2 de membrana. Por lo tanto, la membrana puede separar cargas debidas a 10-12 moles de iones univalentes/volt (la constante de Faraday es 96,516 coulombs/mole de iones univalentes).
El efecto de la resistencia y capacitancia de la membrana se muestra en la Figura 6. El diagrama muestra el arreglo de los experimentos, con un par de electrodos de micropipeta insertados en el axón. Las micropipetas se hacen con capilares de vidrio de 1 a 2 mm de diámetro, estirados hasta formar una punta muy fina (menor de 1 µm en diámetro) y se llenan con una solución conductora, como KCl. En cada una de las micropipetas se inserta un alambre fino y una de ellas se conecta a un estimulador que produce un pulso cuadrado de corriente, mientras la otra se conecta a la entrada de un amplificador, A. La otra entrada del amplificador (recordemos que debemos medir el voltaje entre dos puntos en el espacio) hace contacto con el líquido extracelular y el voltaje transmembrana es amplificado y puede ser visto en un osciloscopio, M.
Cuando se inyecta en un axón un pulso cuadrado de corriente saliente, como el mostrado en el trazo superior (marcado 'corriente inyectada') desde el microelectrodo a la derecha, el sistema de registro ve un voltaje como el mostrado en el segundo trazo. Este voltaje aumenta mas lentamente que la corriente, hasta que alcanza un máximo y después, cuando la corriente cesa, el voltaje cae lentamente a su valor inicial. Entre mayor sea la distancia entre los electrodos de estimulación y de registro (como se indica a la derecha de cada trazo), el voltaje se elevará y caerá mas lentamente, y el valor máximo será menor (trazos inferiores en la Fig 6). Este experimento ilustra porque la constante de tiempo de la membrana debe medirse en el sitio de la inyección, ya que los valores en otros lugares serán imprecisos por la influencia de la longitud de la membrana. Los valores para τ en las neuronas están entre 0.5-5.0 msec.