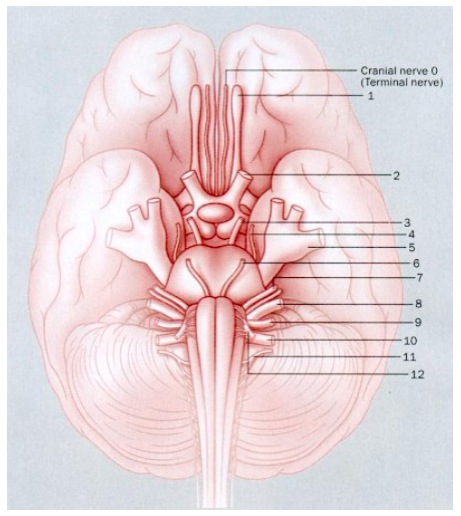
Fig. 1. Esquema del cerebro y la posición de los nervios craneales
El organismo humano es sumamente complejo y es notable que a pesar de deteriorarse con los años, en algunos casos sigue funcionando bien durante aproximadamente 100 años, lo que es mas de lo que vive la enorme mayoría de los animales. Sin embargo, todavía mas notable es que dentro de esa enorme complejidad del organismo, hay un órgano que sobresale precisamente por ser mas complejo que los demás y ese es el cerebro, de quien se dice que con sus aproximadamente 100 mil millones de neuronas y un millón de millones de conexiones, es la estructura mas compleja que existe en el universo conocido.
El cerebro forma parte del Sistema Nervioso y en los humanos tiene tres funciones. La primera de ellas es controlar y coordinar los otros órganos del cuerpo, lo que le permite mantener saludable el organismo mientras efectúa la segunda función, relacionarnos en forma armónica con el medio ambiente. Sin embargo, es su tercera función la que nosotros consideramos mas importante, porque nos hace diferentes a los otros animales y se debe a que residimos en el cerebro; esto es, nosotros, lo que llamamos el 'yo', se encuentra en el cerebro.
Para el estudiante de medicina y el médico el cerebro es también un órgano único, ya que a diferencia de los otros órganos del cuerpo, no sólo sufre alteraciones orgánicas (neurológicas), sino que también tiene problemas cuya base biológica apenas empezamos a vislumbrar y que llamamos mentales (psíquicos).
El conocimiento del cerebro humano avanza lentamente, en parte porque generalmente sólo se estudia en forma directa en personas que han muerto y aunque eso ha proporcionado una gran cantidad de conocimientos sobre las bases de varios problemas médicos, no ha facilitado el conocimiento anatómico o fisiológico de regiones poco conocidas. Por ello, se tiene que recurrir al estudio del cerebro de otros animales y es común encontrar en ellos características que después pueden ser buscadas y frecuentemente encontradas en los nuestros. Esto no es extraordinario, si aceptamos que el cerebro ha estado desarrollándose a lo largo de millones de años de evolución y que somos los herederos de características morfológicas y funcionales que fueron probadas en múltiples cerebros animales antes de llegar al nuestro.
Sin embargo, hay ejemplos en los a pesar que un conocimiento es relativamente antiguo e importante todavía no ha llegado a los libros de texto, como el número de los llamados 'pares craneales'. El número de pares craneales descritos en los libros de texto y por ello conocidos por todos los estudiantes de medicina es de 12, numerados a partir de la región frontal del cerebro. El par número I es el olfatorio, el II el óptico y así sucesivamente. Sin embargo, desde 1878 se sabe que existe otro par craneal adelante del olfatorio, pero como ya se le había adjudicado a este el número I, el nuevo par tuvo que ser llamado par craneal 0 o terminal. Este par craneal 0 fue descubierto por Gustav Fritsch en el cerebro del tiburón y fue hasta 1913 que se encontró en el cerebro humano. Su importancia deriva de que aunque también se origina en los receptores nasales, como el par olfatorio, en vez de mediar la olfación de compuestos químicos en general, está involucrado en la detección de feromonas sexuales, que son mezclas de moléculas que inician conductas sexuales tanto en animales como en humanos y de las que depende la preservación de la especie.
Fig. 1. Esquema del cerebro y la posición de los nervios craneales
Otro conocimiento importante adquirido durante los últimos años del siglo XX y que apenas empieza a ser descrito en algunos libros de texto, se basa en que los órganos abdominales están inervados por plexos conocidos desde hace muchos años, pero que se habían visto como porciones del Sistema Nervioso Autonómico. Sin embargo, estudios mas recientes han mostrado que esos plexos pueden detectar el estado funcional de los órganos abdominales y regularlos en forma independiente del Sistema Nervioso Autónomo. Esto, aunado al hecho de que contienen tantas neuronas como las que hay en la médula espinal, ha originado la hipótesis de que ahí se encuentra un Sistema Nervioso tan completo que ha sido llamado Entérico y al que se le adjudica no sólo la detección y control del estado funcional de los órganos abdominales, sino la capacidad de tomar decisiones en forma autónoma. Como esta característica es una de las que definen un cerebro, empezamos a preguntarnos si esos plexos deben considerarse cerebros y si es así, cuáles pueden ser sus límites.
Durante los primeros pasos de la evolución de los organismos, que se iniciaron a partir de células aisladas, pronto empezaron a hacerse mas complejos y a desarrollar los órganos del cuerpo. Por ello, fue necesario un mecanismo que no sólo sirviera para controlar esos órganos en forma individual, sino también para coordinarlos, ya que sus funciones tenían que ser sincrónicas. Ese papel fue tomado por un grupo de células que derivó en lo que ahora llamamos Sistema Nervioso, por lo que no es sorpresa que haya aparecido temprano en la evolución de los organismos y para explicarlo se han desarrollado varios 'escenarios'.
El escenario mas favorecido actualmente para explicar el desarrollo del Sistema Nervioso se remonta a los animales llamados esnidarios, entre los cuales se encuentran las 'aguas malas', anémonas, corales y otros. La implicación es que en esos animales se desarrollaron células mecanoreceptoras que pueden haber derivado de células 'neuroepiteliales' ancestrales. Como estas células son receptoras y efectoras, pueden detectar estímulos químicos y mecánicos, transducir esas señales a un tipo de energía común a ellas y reaccionar descargando en forma completamente automática una especie de dardo, llamado nematocisto. En una siguiente etapa evolutiva, las mismas células pueden haberse diferenciado en células neuronales, con prolongaciones que empezaron a establecer conexiones (sinápsis) con otras células, por ejemplo células mioepiteliales, involucrándolas en la respuesta al estímulo. Progresivamente esas células neuronales se conectaron con otras células iguales y se empezaron a organizar laas redes, como el anillo nervioso que se localiza en la base de los tentáculos de la cabeza de los esnidarios y que tiene la función de coordinarlos.
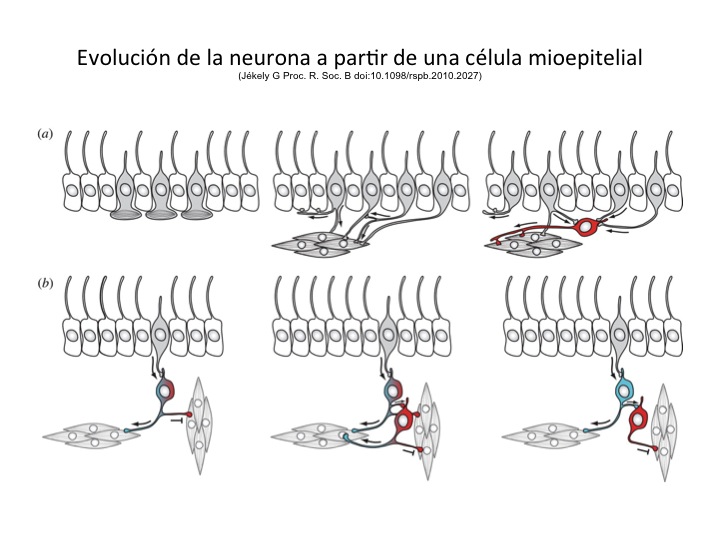
Fig. 2. La hipótesis mas común sobre la evolución de las neuronas
Sin embargo, la característica mas importante de una neurona es que no sólo se divide para pasar a sus descendientes y a través del DNA sus propias características morfológicas y funcionales, sino que también es capaz de crear memorias y planes, y de comunicar esa información a otras neuronas. Esto fue posible cuando las neuronas finalmente desarrollaron dendritas y axones, estructuras especializadas en la recepción y trasmisión de mensajes eléctricos y químicos, que las hacen capaces de comunicar información mas específica y diferenciada.
A lo largo de la evolución el número de células nerviosas secretoras y trasmisoras aumentó en las formas de vida más complejas y el resultado fue que estos organismos tuvieron que aprender a controlar, coordinar y dirigir su comportamiento en una forma mas sofisticada, y sus diferentes partes empezaron a comunicarse por medio de redes neuronales. Estas redes no tenían un centro especializado en recibir señales y mandar instrucciones a otras partes del cuerpo, sino que cada región era autónoma, lo que no era apropiado para conductas generalizadas. Sin embargo, las neuronas pronto formaron un centro especializado en recibir los mensajes de los receptores periféricos, analizarlos y después mandar señales a células musculares para producir el comportamiento apropiado a la condición estimulante.
Los primeros sistemas nerviosos eran redes cuyas neuronas se activaban en forma mas o menos indiscriminada, ya que la excitación de una se extendía por toda la red. En una etapa posterior aparecieron animales alargados que tenían un extremo anterior y uno posterior; esto es, una cabeza y una cola, con lo que la red evolucionó para formar una cadena nerviosa longitudinal, como la que aún se ve en los gusanos y algunos artrópodos. La diferenciación del extremo anterior de la cadena nerviosa continuó con un engrosamiento, donde se concentraron las neuronas para formar un ganglio que empezó a recibir la información proveniente de los órganos de los sentidos localizados en la parte frontal del animal y que es llamado cefálico. Para el control del cuerpo se desarrollaron varias cadenas longitudinales, quizá hasta ocho, que se originaban en el ganglio cefálico y recorrían toda la longitud del animal, con ganglios que tenían las neuronas que se encargaban del control local de cada segmento. Estas cadenas se conectaban entrte si por medio de prolongaciones llamadas comisuras.
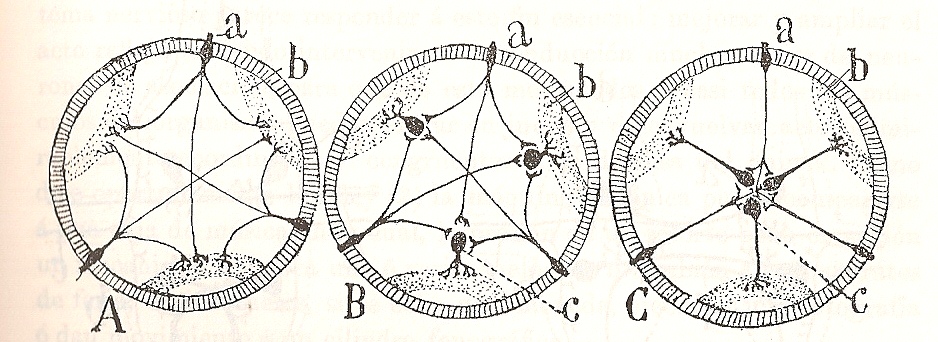
Figura 3. "Esquema mostrando la utilidad de la multiplicación de neuronas y la concentración de estas en ganglios. A, invertebrado ideal donde se supone existen solamente las neuronas cutáneas o sensitivas; B, organismo (actinia?) en el cual existen ya las dos especies nerviosas motoras y sensitivas, pero en el cual no se ha operado aún la concentración de aquellas; C, invertebrado en el cual (vermes) las neuronas motrices se han concentrado en ganglios." (Cajal, 1909)
La diferenciación del extremo anterior de la cadena nerviosa continuó y eventualmente se formó una colección de núcleos y los varios lóbulos del cerebro. Los primeros dos lóbulos fueron el resultado de ganglios de células con características similares que servían dos funciones vitales, un lóbulo olfatorio para analizar información química y un lóbulo óptico para analizar información visual. La expansión de conexiones de estos dos lóbulos llevó a la formación del cerebro moderno.
A partir del lóbulo olfatorio en los mamíferos se desarrollaron otras regiones, entre ellas el sistema límbico, que analiza funciones básicas para la sobrevivencia, como la alimentación, lucha, escape y copulación. Después, conforme el número de neuronas que analizaban información olfativa aumentaba, se empezaron a formar las primeras capas de la corteza. Este sistema olfativo-límbico eventualmente dió origen a la primera corteza motora y a los dos hemisferios cerebrales, que cubrieron casi completamente el cerebro. Así, mucho del cerebro humano ha evolucionado a partir de ese sistema, un hecho que es muy relevante para entender el comportamiento humano.
A lo largo de la evolución y conforme muchos tipos de animales aparecían, proliferaban y frecuentemente desaparecían, también el cerebro se desarrollaba en respuesta a los estímulos de un medio ambiente cambiante. Los animales que habitaban el planeta hasta la llegada de los mamíferos hace unos 100 millones de años, tenían centros motores corticales y sistemas límbicos con sólo dos capas. Con la llegada de los mamíferos, la corteza se desarrolló hacia afuera, capa por capa, cubriendo el cerebro antiguo, de manera que este 'cerebro nuevo' desarrolló cinco o seis capas de corteza que formaron los hemisferios cerebrales. En esta forma y debido a la superioridad 'intelectual' que esa neocorteza proporcionó a los mamíferos, estos evolucionaron, se multiplicaron y pronto dominaron el planeta.
Después, la neocorteza continuó acumulándose y como ya no había mas espacio para ella dentro del cráneo, tuvo que plegarse y formar las circunvoluciones, que expandieron la superficie del cerebro y permitieron aumentar el número de neuronas y conexiones. Sin embargo, todavía actualmente existen mamíferos con diferentes cantidades de corteza; así, la corteza límbica no es muy diferente entre los mamíferos, pero la neocorteza se expande progresivamente conforme pasamos de los mas simples a los mas complejos, como los primates y humanos.
Nuestra característica de humanos es un gran desarrollo de la neocorteza, con numerosas circunvoluciones que acomodan la expansión de los lóbulos, particularmente el frontal y el occipital. Aunque los otros primates poseen una neocorteza considerable, no tienen áreas esenciales para funciones sofisticadas como la producción del lenguaje, que ocurre en la circunvolución angular.
En esa forma y con el paso del tiempo, el grupo de neuronas que ahora llamamos cerebro fue adquiriendo otras funciones que derivaron del aumento en la complejidad de los órganos de los animales, que también se estaban haciendo mas y mas complejos. A todo esto se fue añadiendo también un tipo de funciones que llamamos 'emergentes' y que derivan del aumento en el número de neuronas y de la complejidad de sus conexiones, las que aparentemente dieron base a nuestras "funciones superiores", entre las que se encuentra la conciencia, una de las funciones que mas interrogantes han despertado entre los neurocientíficos.
La morfología del cerebro humano se hace más fácil de apreciar si se conoce cómo se desarrolla en el embrión, por lo que en esta sección haremos un breve resúmen de las primeras etapas del desarrollo del sistema nervioso.
El sistema nervioso se inicia como un surco en las células superficiales de la línea media en la parte posterior del embrión. Con el tiempo este surco se va haciendo mas profundo y pronto forma un tubo de paredes gruesas, separado de la superficie y destinado a formar el cerebro y la médula espinal. Conforme el embrión se desarrolla, la parte frontal cerrada del tubo se expande en tres vesículas conectadas que formarán el prosencéfalo (cerebro anterior), el mesencéfalo (cerebro medio) y el rombencéfalo (cerebro posterior) respectivamente (Figura 3a).

Fig. 3. Esquema del desarrollo embriológico del cerebro. a & c, desarrollo inicial; b & d desarrollo posterior. e, vista lateral semitransparente de un cerebro humano adulto.
Posteriormente la vesícula mas anterior, el prosencéfalo, se subdivide en dos vesículas para formar las regiones llamadas telencéfalo en la parte anterior y diencéfalo (tálamo) en la parte posterior. Del telencéfalo se desarrollarán dos protuberancias que inicialmente crecen en dirección anterolateral y eventualmente también en dirección posterolateral, convirtiéndose en los grandes lóbulos del cerebro que cubren el techo del diencéfalo. Estos son los hemisferios cerebrales, dentro de los cuales se forman espacios llamados ventrículos, izquierdo y derecho (primero y segundo) del cerebro.
| División Primaria | Subdivisiones | Partes constituyentes | Cavidades |
| Prosencéfalo | Telencéfalo | Lóbulos olfatorios | Ventrículos laterales |
| Hemisferios cerebrales | Porción rostral del 3er ventrículo | ||
| Diencéfalo | Epitálamo (epífisis) | ||
| Tálamo | La mayor parte del 3er ventrículo | ||
| Hipotálamo | |||
| Infundíbulo | |||
| Mesencéfalo | Mesencéfalo | Lóbulos ópticos (peces, reptiles, aves) | |
| Colículos/Corpora quadrigemini | Acueducto cerebral | ||
| Pedúnculo cerebral | |||
| Rombencéfalo | Metencéfalo | Cerebelo | |
| Puente (mamíferos) | 4o ventrículo | ||
| Mielencéfalo | Médula oblongata | ||
| Médula espinal | Médula espinal | Médula espinal | Canal central |
Después, el techo del diencéfalo se adelgaza, quedando cubierto por vasos sanguíneos que forman el plexo coroide anterior que sobresale hacia la cavidad (el tercer ventrículo del cerebro) y es importante para el intercambio de gases y la nutrición del tejido. Sin embargo, una pequeña porción del diencéfalo permanece como la glándula pineal o epífisis. Mientras tanto el piso del tálamo desarrolla una depresión con forma de embudo llamada infundibulum y parte de su pared se funde con tejidos del stomodaeum que se está desarrollando, para formar la glándula hipófisis o pituitaria.
La invaginación estomodeal (stomodaeum) es un crecimiento ectodérmico que avanza para acercarse al tubo ciego del esófago anterior del tracto digestivo en desarrollo. Conforme crece hacia adentro, pasa por abajo del cerebro en formación y participa en el desarrollo de la hipófisis. Mientras tanto, las paredes del diencéfalo, laterales y posterior al infundibulum, se diferencian en un centro muy importante, el hipotálamo.
El prosencéfalo tiene una asociación funcional con el sentido del olfato, ya que en su parte anterior recibe entradas sensoriales provenientes de los lóbulos olfativos, mientras el mesencéfalo (cerebro medio) está asociado con el procesamiento de las entradas de los ojos. Los axones provenientes de los ojos entran al cerebro a través del diencéfalo, adelante del infundibulum en el quiasma óptico y terminan en el tectum o techo del mesencéfalo.
El 'quiasma' es un entrecruzamiento de axones provenientes de la retina y está relacionado con la percepción de profundidad. Esto ocurre porque el campo visual que ve cada ojo está dividido en dos partes horizontales y la mitad de las entradas a un ojo que provienen de un lado (ipsilateral) continúan hacia ese mismo lado del cerebro, mientras la otra mitad cruza (contralateral) al otro lado del cerebro. El resultado es que cada lado del cerebro puede comparar la información visual obtenida por ambos ojos y el paralaje es usado para crear la percepción de profundidad (para mas información ver Decusación).
El rombencéfalo o cavidad del cerebro posterior constituye el cuarto ventrículo y se desarrolla en el bulbo del cerebro adulto. Un crecimiento dorsal del techo del cerebro posterior, con apariencia de domo y una superficie convoluta, forma el cerebelo, que tiene la función de mantener el equilibrio de la posición (manteniendo a la persona en posición vertical y orientado en el espacio relativo a la gravedad). También funciona para refinar y coordinar los movimientos motores. Por lo tanto, el cerebelo no inicia la actividad motora, sino que la modula y coordina los comandos para los movimientos que se originan en otras regiones del cerebro, siendo extremadamente importante en este papel de regulador del movimiento.
Posterior al cerebelo se encuentra el plexo coroide posterior, con membranas delgadas, pliegues y muy vascularizado, que se forma a partir del techo de la médula, donde se encuentran los núcleos primarios o raíces de los nervios craneales VII a XII. Es la mayor ruta a través de la cual ascienden y descienden las vías de los centros superiores del cerebro. La médula también contiene centros para los reflejos visceral, auditivo y propioceptivo, así como centros reflejos para la respiración y el latido cardíaco.
En la etapa del desarrollo embrionario en la que el tubo neural se ha diferenciado suficiente como para poder reconocer cinco vesículas cerebrales y la médula espinal, su pared es un neuroepitelio simple y todavía no se han generado neuronas, pero esto cambiará pronto y la forma como ocurre nos proporciona conocimientos fundamentales para entender el plan básico del sistema nervioso.
El patrón de generación de las motoneuronas es particularmente claro en la médula espinal, por lo que empezaremos ahí. Vista en un corte transversal (Figura 1) la médula espinal tiene cuatro partes, un techo delgado en la línea media dorsal, una delgada placa en la línea media ventral, y dos paredes laterales. En un periodo específico del desarrollo algunas células neuroepiteliales dejan de dividirse y migran hacia afuera del neuroepitelio (llamado capa ependimal o ventricular) para formar una zona nueva que se conoce como 'capa del manto' del tubo neural. Estas células son las neuronas jóvenes y han sufrido un proceso irreversible de desarrollo llamado 'determinación' y ya no volverán a dividirse.

Figura 1. "Corte esquemático de la médula de un embrión humano en la cuarta semana (según His). A, raíz anterior; B, raíz posterior; C, epéndimo; a, muro epitelial ó placa interna de His; b, neuroblastos motores; c, velo marginal ó substancia blanca primordial; e, manojo oval; d, neuroblastos comisurales; f, comisura anterior." (Cajal, 1909)
La capa del manto en las neuronas jóvenes se encuentra colocada entre las otras dos capas. La mas interna se denomina 'capa ependimal' y tiene una apariencia pseudoestratificada, cubriendo el centro del tubo neural que se encuentra lleno de líquido y en el adulto será el sistema ventricular. En cambio, la capa externa es una zona llamada capa marginal, que contiene las prolongaciones de varios tipos de células.
En la Figura 1, tomada de Cajal (1909), es claro que en esta etapa del desarrollo la capa del manto es mas gruesa en su parte ventral y gradualmente se adelgaza en dirección dorsal. De hecho, la neurogénesis se inicia ventralmente y se mueve dorsalmente en forma gradual, produciendo un gradiente de generación neuronal de dirección ventral a dorsal en la médula espinal y también en el cerebro posterior. Como resultado, en la región ventral la capa ependimal es delgada, mientras la capa del manto es mas gruesa, un arreglo que resulta en un surco poco profundo que aparece en la pared interna del tubo neural y en forma aproximada divide el tubo en una placa ventral basal y una dorsal alar.
El significado fundamental de las placas basal y alar es que las primeras neuronas generadas en el tubo neural son motoneuronas y sus axones salen en los haces que llamamos raíces ventrales. En contraste, los axones de las neuronas sensoriales en el ganglio de la raíz dorsal crecen en la placa alar, cuyas neuronas no extienden axones en las raíces ventrales. Por lo tanto, la placa basal primitiva está asociada con el sistema motor, mientras la placa alar primitiva está asociada con el sistema sensorial.
La distinción embriológica clara entre la placa basal/raíz ventral y la placa alar/raíz dorsal y sus asociaciones obvias con los sistemas funcionales motor y sensorial respectivamente, confirmó la llamada ley de Bell/Magendie. Esta ley indica que las raices dorsales de la médula espinal llevan información sensorial, mientras las raíces ventrales trasmiten información motora, lo que sugiere una 'circulación' de la información nerviosa. Esto fue fundamental para la idea que gradualmente explicó el concepto del arco reflejo.
Cajal, S.R. Textura del Sistema Nervioso del hombre y de los Vertebrados. Tomo I. 1909. Figs. 3 & 184.